Let us begin with four videos on CRISPER-Cas9 on genome editing technology as a prelude to this post.
https://www.google.com/search?q=a+YOU+TUBE+VIDEO+ON+CRISPER-CAS+Human+genome+editing&rlz=1C1CHZN_enUS948US948&oq=a+YOU+TUBE+VIDEO+ON+CRISPERCAS+Human+genome+editing&aqs=chrome..69i57j69i64l3.33939j0j15&sourceid=chrome&ie=UTF-8#fpstate=ive&vld=cid:2f69e609,vid:ce7rhjSLIWc
https://www.google.com/search?q=a+YOU+TUBE+VIDEO+ON+CRISPER-CAS+Human+genome+editing&rlz=1C1CHZN_enUS948US948&oq=a+YOU+TUBE+VIDEO+ON+CRISPERCAS+Human+genome+editing&aqs=chrome..69i57j69i64l3.33939j0j15&sourceid=chrome&ie=UTF-8#fpstate=ive&vld=cid:eda92929,vid:_PU5bCflzEM
https://www.google.com/search?q=a+YOU+TUBE+VIDEO+ON+CRISPER-CAS+Human+genome+editing&rlz=1C1CHZN_enUS948US948&oq=a+YOU+TUBE+VIDEO+ON+CRISPERCAS+Human+genome+editing&aqs=chrome..69i57j69i64l3.33939j0j15&sourceid=chrome&ie=UTF-8
https://www.google.com/search?q=a+YOU+TUBE+VIDEO+ON+CRISPER-CAS+Human+genome+editing&rlz=1C1CHZN_enUS948US948&oq=a+YOU+TUBE+VIDEO+ON+CRISPERCAS+Human+genome+editing&aqs=chrome..69i57j69i64l3.33939j0j15&sourceid=chrome&ie=UTF-8#fpstate=ive&vld=cid:1c01f789,vid:E8vi_PdGrKg
ABSTRACT
Biology today has invented a new Genetic technology CRISPR-Cas9 that comes to us from our old friends, prokaryotic bacteria and archaea. Amazing is the explicit molecular intelligence involving two RNA molecules, a multifunctional nuclease enzyme that binds to them, and a viral DNA in case of bacterial defense that needs to be snipped out and integrated into the CRISPER locus of the bacterial genome shown diagramiticaly below. The technology evolved in less than two decades dealing with a double RNA guided nuclease enzyme that breaks the double strand of DNA and helps integrating the snipped out viral segment as spacer in the CRISPER locus of the bacterium. The two RNAs are crRNA and trcrRNA. They can be fused into one SgRNA for a simpler operation also The cell repairs the break naturally after deletions or insertions are made. Thus we have a new DNA. We call this rewriting of genetic code “gene editing”.
The Khan Academy
The endonuclease Cas9 is a dual RNA-guided enzyme associated with the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) now confirmed as the adaptive immune system in Streptococcus pyogenes. It is now designed as sgRNA complex (crRNA and tracrRNA put together). The complex uses PAM existing in human genome. Dual RNA (crRNA and tracrRNA) molecules or a fused version thereof guides CRISPR-Cas9 system to a DNA locus to be chopped past a few nucleotides from marker PAM. Please review the video as to its structure.
https://www.google.com/search?q=youtube+video+on+Cas9+crystal+structure&rlz=1C1CHZN_enUS948US948&oq=youtube+video&aqs=chrome.0.69i59j69i57j35i39j0i131i433i512j0i20i263i512j0i131i433i512l2j0i512j0i131i433i512j0i271.14996j0j15&sourceid=chrome&ie=UTF-8#fpstate=ive&vld=cid:76c3e64d,vid:7C20s4eAGtU
The old bacteria have come to our help and the Cas9 nuclease from Streptococcus pyogenes with customized and synthetic single guide sgRNA using the PAM (protospacer Adjacent Motif) are now being routinely used for correcting gene mutations that cause diseases. The principle behind their CRISPER immune system allows us to create new drugs, new plants and crops, and new pets. Let us hope that our ethical standards would keep us away from designing human babies and create havoc in society.
This post is for public education and this is the reason why I have added appendices on CRISPER-Cas classification, CRISPER-Cas vocabulary, and a chronology of research that lead us to this gene editing tool. Our readers can consult the appendices as and when necessary while reviewing this post. Even more important, I think, is incorporating a section on “The Central Dogma of Omics in Molecular Biology” that connects DNA to RNA to proteins, and finally to metabolites.
For editing to proceed, there has to be a way to cut two strands of DNA and the scientists have found Cas9 nuclease to be safe, simpler and accurate compared to TALEN (Transcription Activation Like Effector Nuclease or the Zinc Finger Nuclease (finger-fold zinc ion mediated protein). Double strand break involving the cleavage of the phosphate-sugar backbone on strands of DNA at the same time is necessary. Recent research (https://www.biorxiv.org/content/10.1101/2022.06.13.495754v1.full.pdf) on binary, ternary and quaternary structure of CRISPER-Cas 12a2 nuclease shows that it could be even more powerful editing tool utilizing a single strand RNA, single strand DNA, and double strand DNA breakage indiscriminately. Class II type V CRISPR-Cas12a is a new RNA guided endonuclease that has been recently harnessed as a more powerful Swiss knife like DNA chopping and editing tool.

Genetic Engineering and Biotechnology News, Jan 5, 2023
In this illustration based on cryo-electron microscope images, a Cas12a2 protein unzips a DNA double helix, allowing it to cut the single strands of DNA (blue and green). [Jack Bravo/University of Texas at Austin]
https://www.google.com/search?q=youtube+on+the+structure+of+Cas12a&rlz=1C1CHZN_enUS948US948&oq=youtube+on+the+structure+of+Cas12a&aqs=chrome..69i57j33i160.18454j0j15&sourceid=chrome&ie=UTF-8#fpstate=ive&vld=cid:277caa50,vid:DtWOLG1KCqA
INTRODUCTION
First of all let us get a clear understanding of the constant length CRISPR array of a dyad symmetry. First discovered in E. coli 30 years ago, the conserved repeats of 21-40 nt length exhibit very little variation in sequence. They have been found to be conserved in all archaea and half of bacteria. The constant length spacers are non-conserved sequences. They are homologous to the pro-bateriophage, bacteriophage, and plasmid DNA. CRISPR repeats and spacers are located at intergenic regions of DNA and there is always a common several hundred nt long leader on one side of the repeat cluster. CRISPR array is not present in eukaryotes.
There are four different associated nuclease genes found adjacent to CRISPR genes. “J. Bacteriology, April 1, 2018, volume 20, issue i” is a very good reference to consult. CRISPR represents a mobile genetic element called CASPOSONS.
The figure below gives examples of E. coli and archaea H. mediterranei. Along with CRISPR, we should also get an understanding of Transporons and casposons.
A casposon is a member of a distinct superfamily of archaeal and bacterial self-synthesizing transposons that employ a recombinase (casposase) homologous to the Cas1 endonuclease. They appear to have given rise to the adaptation module of CRISPR-Cas systems as well as the CRISPR repeats themselves. The transporons are defined as DNA segments that can undergo transposition between chromosomes, phage, and plasmid DNA in the absence of a complementary sequence.

https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5847661/figure/F1/
This primitive system has now become a hot topic in synthetic biology directed to gene editing and creating new genes or modified genes.
PUBLIC EDUCATIONS ABOUT GENES AND GENOME
The objective of this post is public education about genetics and about who we are. Whereas genomics deals with the entirety of genes and their interactions as to complex diseases, genetics deal with individual genes as the units of heredity. There are diseases like sickle cell anemia, cystic fibrosis, phenylketonuria, and Huntington’s disease due to aberrations in a single gene. Genes are made of DNA, the universal information molecule for all that lives and modification of the genome is tantamount to modifying the millions of years old aggregate of the evolutionary information. The public wins by learning as much as it can about genetics and genes. The public needs to know that biomolecules that make and run our lives are super-intelligent.
THE CENTRAL DOGMA OF MOLECULAR BIOLOGY: DNA, GENES, AND CHROMOSOMES
(Precis Clin Med. 2021 Sep; 4(3): 179–191)

https://www.shutterstock.com/search/central-dogma
Whereas only 0.5 % of our body weight is DNA, RNA, the work horse, makes more than 5% of our body weight. Topology of DNA and RNA molecules is the key to their molecular functions. With a width of 2 nanometers, the two strands of DNA turn righthanded or clockwise every 10 base pairs. The double strand of DNA whose lower strand complementary to mRNA is the 5’ to 3’ coding SENSE strand. The other Antisense strand is mRNA like 3’ to 5’ strand. The numbers refer to carbon number of ribose sugar to which phosphate group is attached. The phosphodiester bond is between 5′ and 3′ carbons of ribose molecules. The complementarity of two strands ensures hydrogen bonding between pyrimidine (T, C) and purine (A and G) nucleotides and permits helix formation. Whereas prokaryotes like E. coli and Streptococcus pyogenes have single DANA genome without nucleus, the eukaryotes including humans have a much larger 3.5 billion base pairs genome of 23 chromosomes pairs wrapped around a histone protein structure within a nuclear membrane. The bacterial DNA is at the most 8 million bases long and can encode up to 6000 genes. The human genome encodes 20,000 to 25000 genes. In case of humans, mitochondria has a 16,500 nt long circular chromosome also capable of making all proteins for oxidative phosphorylation via electron transport chain, 22 transfer RNAs, and two ribosomal RNAs. Now you can see how friendly the bacteria are to us because they made our mitochondria, they inhabit in our intestine, and now they are helping is edit our genes for a disease free life.
DNA is a rigid polymer of 23.7 A width and 34 A per 10 bp length and yet capable of bending, twisting, and compressing. The width of major and minor groves are 22 A and 12 A respectively. Both double stranded DNA and single strand RNA molecules can configure themselves and create a specific 3-D structure with motifs. There can result quadruplexes and coaxial stacks. The most common unmethylated B-DNA has the least twist from the axis, A-DNA has a wider groove and tilt, and Z-DNA is lefthanded zigzag helix. In the absence of any proteins bound to it, it can supercoil and collapse to itself. It can configure into catenanes (inseparable links) and knots. DNA is a unique molecule that provides for mechanisms of copying, coding for 20 amino acids by base triplets, and binding with proteins. Whereas DNA, the deoxyribonucleic acid has deoxy ribose in its nucleotides (purine or pyrimidine boned to ribose sugar which is bonded to phosphate group), RNA has ribose. The base pair in either affords the hydrogen bonds. For example a G-C pair affords 3 hydrogen bonds and an A-T pair affords only two hydrogen bonds. Examine the following pictures for details of 5′ to 3 linear chain of nucleotide (sugar bound tp phosphate and either pyrimidine or a purine) and the steps of pyrimidine-purine bases as rungs of a ladder.

https://bio.libretexts.org/Courses/Portland_Community_College/Cascade_Microbiology/22%3A_Appendix_B_-_Molecular_Genetics_Review/22.2%3A_Structure_and_Function_of_DNA
A sequence of three nucleotides together in a gene encodes an amino acids. A larger portion of sequence deals with where, when, and how a protein is made. An average coding gene can be 3000 to 2.3 million letters (bases) long, the shortest being 500 letters long. Imagine titin muscle protein with a molecular weight of 3 million made of 26,000 amino acids that will require mRNA codon letters of 81,000 and then we have collagen Type I as most abundant protein in our body with only 19 amino acids. And then imagine the size of corresponding mRNA molecules for synthesis of the large and small proteins with the following specifications.
1. A DNA molecule has promoter regions upstream of a gene where relevant proteins (RNA polymerase, transcription factors) are activated for transcription. DNA molecules bend so that enhancers and promoters can be in close proximity for transcription to proceed. A mutation in the promotor region can be just as bad in causing a disease as the one in the gene itself.
2. The upstream switches, regions of DNA that can be bound by a particular activator or repressor in a sequence- specific manner, of a gene that react to external signals in relation to terminating the gene on or off. A switch could be used in combination with many genes. A given gene could be activated by many switches that may be activated by different switch proteins and the genes encoding amino acids for same process have similar switches. A single switch protein may activate multiple genes. Regulatory molecules bind to the switches and recruit RNA polymerase to bind to the gene’s promoter region, increasing the transcription of the gene into messenger RNA. The enormous complexity of gene regulation in higher organisms, combined with the packaging of their DNA into chromatin has yet to be fully understood.
3. There exist different combinations of exons for different mRNAs for the same gene.
4. On 5′ and 3′ ends, there are 100 to 1000 untranslated regions that help attach or detach proteins. The mRNA is by design kept in definite area of the cell.
5. A mRNA molecule has start and stop codons for protein synthesis.
6. More than one genes often act as operon that exist adjacent to one another for transcibing a single promoter.
The 20,000 genes are only 2% all nucleotides in human genome. Part of the genome, it should be pointed out, encodes rRNA and tRNA. The rest is for structural support. 2600 genes code proteins that bind to DNA and the proteins that activate the switches. Exons are sequences present in mRNA, some of which may encode amino acids in a protein. And then there are START CODONS in mRNA for methionine as the first amino acid in all proteins which often drops off later. Also, there are gene introns that do not remain with the mature mRNA following transcription of a gene. For terminating the addition of amino acids, there are stop codons. Our life processes thus depend on the structural complexities designed for the specific functions of nucleotide sequences of DNA and the transcript of its sense strand or mRNA.
mRNA can encode amino acids, act like an enzyme for catalysis, and transmit genetic information. iI’s secondary structure is a hairpin loop type. In eukaryotic cells, the 3′ end of mRNA is a non coding polyadenilate. By convention, 5′ end is the cap and 3′ end is the tail. mRNA is made in nucleus but functions with ribosome outside nucleus in the cytoplasm. There are 2 to 3 mRNAs for every gene and this number depends on the state of growth of the cell. The number of ribosomes is 10 million in in an eukayotic cell. The ribosome complex can join 200 amino acids per minute but even at this rate putting together a molecule of titin nuscle protein of 30,000 amino acids can take up to 3 hours. A rapidly growing mammalian cell may have up to 10 million ribosomes, free or bound to the endoplasmic reticulum. The process of translation on demand and in time of 10 billion proteins in a human body by mRNA molecules that act as coding agent for 20000 genes is a miracle of molecular intelligence.
The strands grow on the 5′ side during the synthesis of DNA. Shown below in the diagram are the nucleotide base, sugar (blue), and phosphate (yellow). Please note that the ribose sugar on the right strand is inverted on its complementary strand. As the new nucleotides line up opposite each parent strand by hydrogen bonding, enzymes called DNA polymerases join the nucleotides by way of phosphodiester bonds between C5 carbon of one sugar C3 of the other as the chain of strand lengthens. DNA polymerase III is a tripartite protein machine responsible for replication of bacterial genome.
![meet 일반생물학] 분자생물학 8-3. DNA 복제 - leading strand와 laggin strand / 각종 효소의 작용 : 네이버 블로그](https://i0.wp.com/mblogthumb-phinf.pstatic.net/20121220_291/dyner_1355981380737aR1PA_JPEG/chain_elongation.jpg?resize=707%2C478&ssl=1)
OMICS dictates all life whereby the 5’ to 3’ sense strand is transcribed into mRNA and mRNA and tRNA helps translate genetic codes into proteins by ribosomes. The proteins run life as inputs to metabolomics. DNA transcribes into mRNA, a molecule that acts like a road on which travels the ribosome code by code. tRNA brings the amino acids to the ribosome site for making proteins. mRNA allows translation of DNA codes into proteins and the proteins as enzymes, neurotransmitters, or transcription factors run the metabolism. In the omics processes, information flows from DNA → RNA → protein, a directional relationship known as the central dogma of molecular biology. It is believed that RNA predates, DNA and Protein. There is an army of mRNAs in a cell depending on number of protein to be synthesized. A single mRNA can encodes a single protein. On the other hand tRNA is specific to each amino acid. The figure below details the omics processes.

https://www.majordifferences.com/2022/04/difference-between-genome-transcriptome.html
DNA is double-stranded, but only one strand serves as a template for transcription at any given time. This template strand is called the noncoding strand. The nontemplate strand is referred to as the coding strand because its sequence will be the same as that of the new RNA molecule.
A ribosome (dark brown) travels along mRNA (reads sequence of nucleotides) as protein synthesis continues and amino acid specific tRNA brings amino acids by the codon. The ride continues until the protein is completely synthesized. The cell has to assemble mRNA, ribosome, tRNA, and amino acids in order to begin transcription or building of proteins from mRNA.
https://passel2.unl.edu/view/lesson/3ccee8500ac8/5
Let us examine in detail the diagram below. On the left is shown the making of mRNA within the nucleus. mRNA transfers to the cyroplasm. It is shown as the code carrying long string of nucleotides. Ribosome rides on mRNA code by codes and the tRNA brings an amino acid, and the protein chain lengthens by additional peptide bond formations. mRNA can be translated by several ribosomes at the same time.
![]()
https://www.microbe.net/wp-content/uploads/2011/05/translation.jpg

http://bscb.org/wp-content/uploads/2014/02/schematic.jpg
Let us repeat and reemphasize the totality of omics processes. DNA, the information molecule of life, first transfers the information to mRNA by a process of transcription. The lower strand of DNA is transcribed into mRNA. The net process of translation takes place at the ribosome that travels along mRNA codes to successively assemble proteins with the help of tRNA. Proteins, as products translation, are molecular machines for life of the living. The genetic code is read by the complex “protein -rRNA” structure of ribosome above. To be a bit more specific, the ribosome as a 20-30 nanometer protein molecule in diameter can deal with three tRNAs at the same time including one whose amino acid has been added, another whose amino acid is being add, and the third who is standing in line with another amino acid.
Please pay attention to yet another example of molecular intelligence. Ribosomal RNA (rRNA) combines with proteins and enzymes in the cytoplasm in order to form the complete structure of ribosome. The ribosome bound rRNA base pairs with tRNA and tRNA binds to the anticodon of the mRNA. The nucleotide sequence of mRNA spells the code of amino acids in a protein. Ribosomes, a kind of rRNA ribozyme, can travel along the length of mRNA starting often with its first codon AUG for methionine. tRNA brings amino acids to the ribosome for elongation of the protein chain until the process reaches to an anticodon on the mRNA. tRNA is small 76 to 93 nucleotide long molecule acylated to a cognate amino acid. Its secondary structure is like a clover leaf and the tertiary structure is L-shaped. tRNA is an adapter between mRNA and the protein being synthesized. Its one end binds to a specific amino acid and the other to the specific codon of mRNA. Cod AUG for methionine is the point of initiation at the 5’ end of the mRNA which is capped with guanidine phosphate for recognition. Ribosomes can join 200 amino acids per minutes. The novelty of CRISPR-Cas9 gene editing tool lies in making or not making proteins at will.
A copy of our genome of 3.2 billion bases is present in all our 32.7 trillion cells of 300 types. DNA is made of chemicals Adenine (A), Cytosine (C), Guanine (G), and Thymine (T). Adenine pairs with Thymine on the second strand and G pairs with C. Three of them make a code for an amino acid.
Chromosomes are made up of a DNA-protein complex called chromatin that is organized into subunits called nucleosomes. The strands of DNA are coiled up in chromosomes present in the nucleus of cells. A section of long piece of DNA in the chromosome makes a gene, the recipe for making a protein. There are 23 pairs of chromosomes: numbers 1 to 22, and the 23rd which is XX in girls and XY in boys. Each chromosome has many genes. Out of 200,000 genes in the genome, a particular cell chooses the genes it wants “turned on” or “Turned off”. That is how the heart, brain, liver, eyes, skin, kidney, and lung cells function differently. Cells need to make thousands of different proteins in the right amounts, at the right time, and in the right place in order to work properly. There are a few exceptions though.
1. Mature red blood cells have no DNA
2. Gametes (sperm and egg cells) have only half of the amount of DNA
3. B cells in our immune system have some DNA that is rearranged in order to make antibodies. Human B cells can produce up to 10 exp 12 antibodies. This is more than the number of genes in the genome. Our cells can rearrange gene segments in order for it to act like another gene. This is the best example of intelligence of DNA segments.
Proteomics studies the dynamics of protein production and their interactions. Metabolomics is also an intermediate step in understanding an organism’s entire metabolism. The amino acid codes of triplet nucleotides are the same for all cells. The cell cytoplasm should have ample supply of amino acids, sugars, and nucleotides as monomer substrates. Here is a Table of nucleotide triplet codes or codons for amino acids written from 5’ to 3’ end with methionine as a start signal.
https://passel2.unl.edu/view/lesson/3ccee8500ac8/6
Although the amino acids are specified by each mRNA codon and multiple codons may code for the same amino acid. Although methionine is the first amino acid incorporated in all proteins made at the ribosome, it may not be there in mature proteins. Please see below an example of genes as DNA segments that make four different proteins: Hemoglobin that carries oxygen, Opsin that lets us see colors, and amylase that digests starch. There are different genes for each one of these protein products.

https://www.thetech.org/ask-a-geneticist/ask293
RNA molecules are the work horses of our cells. They are the real factory workers. mRNA or messenger RNA carries the genetic information out of the nucleus to the cytoplasm. rRNA is like a machinist that helps make the protein including the formation the peptide bonds. tRNA is in the cytoplasm. Its anticodon end binds the mRNA and the other end is amino acid specific. There are twenty tRNAs, one for each amino acid. There are other RNA types involved in structural, catalytic, and regulatory functions (examples are helicase, dicer, and siRNAs). mRNA, rRNA and tRNA are common to both prokaryotic and eukaryotic systems. RNA performs many functions in an organism such as coding, decoding, regulating, and expressing genes. RNA is found in both the nucleus and cytoplasm of humans cells.
SCIENCE OF PASSIVE BACTERIAL IMMUNITY
The world wide scientific community did enormously dedicated research and created more than 1400 patents just in a decade on CRISPR-Cas systems just to find and understand the mechanism of passive immunity in bacteria and archaea.

https://www.fdli.org/2021/11/the-future-of-food-crispr-edited-agriculture/
Moreover, only in two decades the scientists found that bacteria and archaea has evolved for using and manipulating Omics (transcription of a strand of DNA to mRNA and translation of mRNA codons to a complete protein) for their survival by creating an immunological defense. As I begin to trace and present this molecular intelligence explicit in the CRISPER-Cas systems, I recollect India’s Rishi Kashyap’s Vedic Vaiseshika for its reliance on direct observations and inferences that emerge from atoms and molecules. Our modern day scientists did exactly that. They learned by direct observation and inference that bacteria and archaea that evolved 3.5 billion years ago had learned how to re-engineer their genome by snatching away part of the DNA sequence of their viral predators and integrating it into their own DNA. Their ability to do so depended on information and intelligence of the molecules of DNA, RNA, and proteins.
What an exquisite chemistry it is! An RNA molecule becomes part of the Cas protein (a multifunctional nuclease enzyme) and guides the Cas9 system to DNA locus to be chopped past a definite 2-3 sequence of nucleotides from the marker PAM (https://www.microbe.net/wp-content/uploads/2011/05/translation.jpg). This display of molecular intelligence is the story of CRISPER (Clusters of Regularly Interspaced Short Palindromic Repeats)- Cas systems.
First, let us understand the palindromic repeats. A palindromic sequence of DNA is unique in that it can undergo hairpin folding and unfolding (top) and so can its transcript (bottom). This is critical to the Cas9 system’s function. Palindrome is like a word, verse, or sentence (such as madam or number 1881) that reads the same backward or forward. In case of CRISPR the letter is the nucleotide sequence. An example of a palindromic sequence is 5′-GGATCC-3′, which has a complementary strand, 3′-CCTAGG-5′. The CRISPR-Cas locus is the CRISPR array of repeats and spacers and the adjacent Cas genes. Let us note that in 25% cases the Cas genes may be away from the CRISPR array in the chromosome’s DNA. This is the sequence where the restriction endonuclease binds to and cleaves at a specific cleavage site on DNA. Furthermore, the diagram below shows that the spacer (in white) of the CRISPR locus can loop and the repeats can make double stranded stem of a hair pin. This secondary structure is critical to recognition.

https://www.mpg.de/11823627/crispr-cas9-palindromes-structure
Palindromes in Genome (Max Plank-Gessellschaft, MPG Aet for Science).
The picture above represents the palindrome sequence in the DNA of the bacterium Streptococcus agalactiae. Parts of the letter sequence of one strand (green) correspond to those of the other strand (yellow) in the reverse order. However, the palindrome is not perfect. It also contains a non-palindromic sequence (white). The DNA can form hairpin structures using a broken palindrome like this one.
As pointed out earlier, the CRISPER and Cas genes work together and are often located adjacently. The first intelligent system is the CRISPER (Clustered regularly interspaced short palindromic repeats of 24 to 28 bases) array comprised of the repeat nucleotide sequences and the spacers of similar length that represent viral DNA segments snipped out of the DNA of the invading virus. Crispr is transcribed into crRNA/gRNA including the spacer, that guides the Cas nuclease enzyme. It is a kind of 70-80 NT long anti-repeat. The palindrome gene is the key. gRNA or crRNA is complementary to target DNA. tracrRNA that binds to Cas-9 also has short hairpin loops. Cas9 enzyme follows gRNA (crRNA) to DNA that needs to be chopped. sgRNA is crRNA fused to trcrRNA for reasons of simplicity in editing operations. PAM ( Proto-spacer Adjacent Motif) is 3-4 nucleotides upstream the target DNA sequence. Let us note also that the Cas system from different organisms recognize different PAM.
Palindrome sequence in the DNA of the bacterium Streptococcus agalactiae is shown above. Parts of the letter sequence of one strand (green) correspond to those of the other strand (yellow) in the reverse order. However, the palindrome is not perfect. It also contains a non-palindromic sequence (white). The DNA can form hairpin structures using a broken palindrome like this one. The palindrome reads same both forward as backward (https://www.mpg.de/11823627/crispr-cas9-palindromes-structure). Whether DNA or RNA, the hairpin folding is a recognition mechanism. The circular white nucleotides originated with the spacer from the viral genome, two palindromes make the stem of the pin and the three nucleotides make the hairpin arms. The linear strand of RNA becomes a hairpin.
A few noteworthy details are 1. similar nt lengths of repeats and spacers, 2. spacer’s ability to make a circular loop, and 3. the ability of sgRNA to complex with the Cas nuclease. Often crRNA alone can’t do the job and, therefore, the need for tracrRNA which is complementary to the repeats.
Trans-activating CRISPR (tracr) RNA, a small trans-encoded RNA, is a distinct RNA species that interacts with the CRISPR crRNA to form the dual guide gRNA in type II type Cas9 system. It was first reported in Streptococcus pyogenes. The tracrRNA has the recognition sequence necessary for binding by a Cas-9 like DNA nuclease. The tracrRNA and crRNA form a complex, bind to the Cas9 nuclease, and direct the nuclease to the target sequence via the crRNA. The tracrRNA-crRNA interaction is essential for pre-crRNA processing as well as target recognition and cleavage. Pre-crRNA processing by Cas-9 simply makes a smaller guiding RNA.
Cas9 is a single protein that binds to the fused crRNA and tracrRNA as a single guide to target DNA. The cascade below is common to multiunit nuclease of Class I enzyme system.

https://www.neb.com/tools-and-resources/feature-articles/crispr-cas9-and-targeted-genome-editing-a-new-era-in-molecular-biology
Please note the sequence of the repeat-spacer-repeat-spacer array. This diagram shows sgRNA as the guide complementing and recognizing the target DNA segment.

https://www.cell.com/molecular-therapy-family/nucleic-acids/fulltext/S2162-2531%2817%2930058-6
The CRISPR array is transcribed into a long primary pre-crRNA that forms a duplex with tracrRNA containing a complementary sequence. Cas9 specifically promotes pre-crRNA:tracrRNA duplex annealing. The virus invades (top left). It is integrated into crisper locus. CRISPR locus is transcribed into pre-crRNA which is fused into sgRNA that guides and binds to viral DNA on the top right.
Cas9 nuclease anneals crRNA and tracrRNA on its surface and then the dual RNA acts as a single guide for the nuclease to the target DNA.

https://www.thebaileylab.org/posts/crispr-spacers-asymmetry-and-orientation
Pictorially speaking, sg RNA binds to Cas9 which is a ribonucleoprotein complex where the scaffolds of traceRNA bind to the Cas9 protein.

https://www.genepossibilitieshcp.com/approaches/crispr-cas-gene-editing
The picture below, when expanded, clearly shows shows DNA in yellow and RNA in blue.

https://www.vox.com/2018/7/23/17594864/crispr-cas9-gene-editing : The Streptococcus pyogenen’s system Cas9 Nuclease (RED), DNA (yellow), and RNA (blue)
The bacterial immunity system now teaches us how to edit our own genes and those of our animal and crops but with possible problems of writing out large segments of genes and even triggering cancer. We have to perfect this molecularly intelligent system. I am particularly impressed with the possibilities of perfect and flawless food fermentations by lactic bacteria, yeast and Aspergillus oryzae like fungus. Even more important is going to be the hydrogen gas fermentation for the production of more or less pure bacterial proteins like solein (https://www.solein.com/what-is-solein).
We have been using microbes as workers for the production of food and medicine for a long time but using their individual CRISPR-Cas systems of immunity for a large-scale human and plant genome engineering is new and novel. The novelty demands that the CRISPR-Cas knowledge be communicated to public effectively (2,3,6,9, 19, 29, 31, 40, 48, 53, 56). This is what drove me to write this post and discuss the central dogma of genetics, the bacterial CRISPR-Cas passive immunity, and its use as a genetic engineering tool for creating and modifying genes for human therapeutics. Beyond this main focus, I have added to this post three appendices so that the readers have all details on their fingertips as they attempt to master this new science: 1. Cas Classification, 2. CRISPR-Cas Vocabulary, and 3. A chronology of CRISPR-Cas R & D works. I believe that the work in this area will continue during a few more decades and, therefore, everyone on the planet needs to commit to a continuing education on CRISPR-Cas Science and technology. Also, I should advise my reader to listen to TED talk by Dr. Jennifer Doudna (https://www.ted.com/talks/jennifer_doudna_how_crispr_lets_us_edit_our_dna?language=en) and her Nobel Prize acceptance speech (https://www.youtube.com/watch?v=KSrSIErIxMQ) as a primer in addition to four videos included up front on this post.
The big evolutionary mystery is how a complementary RNA finds the target DNA to be chopped, both strands at the same time, just 2 to 3 PAM nucleotides away. Both RNA molecules and Cas protein with their ways of configuring and with their specific topologies have ability for such an intelligence and recognition.
No doubt! The history of palindromic repeat sequences in DNA of bacteria, archaea, and plasmids is unique. The mechanism of RNA guided CRISPR-Cas system has been successfully repurposed now for human gene engineering in less than two decades. CRISPR or Clustered Regularly interspersed Palindromic repeats have now become part of our vocabulary in molcular biology. The history of how bacteria used repeats of nucleotide sequence (CRISPR) to edit their own genes is necessary to appreciate how CRISPR-Casa enriched biology is going to revolutionize personalized medicine, health, and nutrition in very near future. The evolution of repeats and their embedded spacers as information or memory is something we have not yet fully understood.
THE REPEATS IN THE GENOME OF BACTERIA: THE MYSTERY
Only 28 years old Francisco Mojika finds multiple repeats of 30 bases in the DNA of salt tolerant archaea Haloferax mediterranei at the university of Alicante in Spain. The repeats were separated by spacers of 36 bases not known before. Mojika spends a decade demystifying the finding. He found the same pattern in H volcani. Similar work on E coli by Ishino in Japan tempted him to speculate that the repeats are something very important. He goes to Oxford for a Ph.D. and returns home as a faculty member at the University of Alicante. By 2000, he found the repeats in 20 different organisms including Mycobacterium tuberculosis, Clostridium difficile, and the plague bacteria Yersinia pestis. Soon there were reports on functionally related CRISPR Associated nuclease genes in the vicinity of the repeats. But no clue yet as to what is the functional value of the repeats. No doubt! Francisco Mojica is the clear leader in the CRISPR field. Then big-data bioinformatics comes along and does wonders for him.
MORE RESEARCH ON REPEATS ESTABLISHES THE ADAPTIVE IMMUNITY CONCEPT
Busy sequencing E coli gene, Mojika finds that one of its spacers matches the P1 phage sequence that had infected the E coli. This was an eye opening finding. In just one week, he checked 4500 spacers, 88 of which had known sequences. That is two third of the 88 matched viral sequence. Now Mojika has a clue to adaptive immunity. He sent his paper to Nature. Nature rejected the manuscript. PNAS, Molecular Microbiology, and Nucleic Acid Research did exactly the same for presumed lack of novelty. After much frustration the paper on CRISPR’s function on adaptive immunity did appear in J. of Molecular Evolution in 2005. About the same time Gilles Vergnaud of France’s ministry of defense working with Universite Paris-Sud begins work on the plague bacteria Yersinia pestis. His colleague Christine Pourcel finds CRISPR locus in Yersinia pestris with spacers analogous to phage DNA. Coincidently, Vergnaud faced rejection by PNAS, J. of Biotechnology, Nucleic Acid research, and Genome research also trying to publish his findings. Finally, he got it published in Microbiology on March 1, 2005. Alexander Bolotin, a Russian émigré at Frances National Institute of Agricultural Research, also published a CRISPR paper just one month after Mojica’s paper. Bolotin was the first to coin Adaptive Immunity in bacteria against invading viruses.
EXPERIMENTAL EVIDENCE
Then comes experimental evidence by the work by Philip Harvath of Rhodia/Danisco while trying to find phage resistant Streptococcus thermophilus for cheese production. Horvath finds that spacers relate to phage resistance. This work corresponded to the close time line of Mojica and Vergnaud’s publications and is similar to findings by Rodolphe Barrangou of Danisco and Sylvain Moineau of University Laval in Quebec City, Laval. It is confirmed now that the resistant strains had phage derived sequences as spacers in their CRISPR loci. Multiple spacers meant more resistance. Also, they studied Cas7 and Cas9 genes. Cas9 was an active participant in the bacterial immune system which depended on precise DNA sequence match between spacers and the target viral DNA (61,63).
PROGRAMED CRISPR BASED IMMUNITY
CRISPER gets programmed by by John van der Oost of University of Amsterdam with help of his connections at Helsinki, Heidrlberg and finally the Wageningen University where he has to work on extremophiles from hot springs of Yellow Stone Park in USA. He collaborated with Eugene Koonin of National Center for Biotechnology Information (NCBI of NIH). Koonin introduced Van der Oost to CRISPR. They inserted one E coli system to another and examined a number of Cas nucleases. E coli has Class I of a complex cascade. They found that crRNA started with last 8 bases of the repeat sequence followed by a spacer and the beginning of the next repeat region. They imagined that the palindromic natures of repeats permitted a secondary structure to crRNA. They created artificial CRISPER array against four genes of phage lambda. As predicted the strains carrying new CRISPR sequences showed resistance against phage lambda. Now we have a case of programmed CRISPR based immunity, a flue shot for bacteria so to say. CRISPR, they now know targets double strand DNA breakage for fighting against the viruses by steeling their DNA fragment. A few noteworthy points in these regards.
-
The idea that the target is DNA came from Luciano Marraffini’s work on Staph epidedermidis while working with Northwestern University Faculty member Malcom Casadaban. They found that CRISPER blocked plasmids as it did the viruses.
-
Cas9 is guided by crRNAs and creates double stranded DNA Break.
-
Trans-activating crispr RNA (tracrRNA) is a small trans-encoded RNA (60) discovered first by Nobel Laureate to be Emmanuelle Charpentier who studied human pathogen Streptococcus pyogenes. This is the small RNA with short hair-pin secondary structure.
-
Similar immunity mechanisms were found to be reconstituted in Distant Organism (38).
-
CRISPER in vitro for double strand break three nucleotide away from the PAM sequence (27) were now well recognized in that two nuclease domains cut opposite strands.
-
Universal programing by crRNA and tracrRNA acting as a single sg RNA can do double strand break of DNA.
-
Inserting DNA at a specific location in mouse embryonic stem cells and producing a modified mouse was inefficient but double stranded break did the trick using a nuclease. This made homologous recombination simple via zinc finger nuclease. Now Programmable CRISPER came as boon. Fen Zhang who got a Ph.D under Karl Deiss at Stanford found a way when he got to Harvard. He used S. thermophilus Cas9 for use in human cells and engineered crRNA. He later found that Cas9 from S. pyogenes was much better with tracrRNA and CRISPER array. He could mutate genes. He could edit multiple genes simultaneously. This is happening around at the same time when Emmanuelle Charpentier and Jennifer Doudna are publishing their findings at UC Berkeley. Zhang’s 2013 science paper became the most cited of all papers. The learning from the distinguished professor George Church next door paid him off. They could spread synthetic genes through natural populations. They found that 3’ hairpin of tracrRNA became essential for CRISPR-Cas system function.
-
CRISPR goes viral in Yeast (21), nematode, fruit fly, zebrafish, mouse, and monkey. Then talks of potential applications in medicine and agriculture became rampant. The talks of designer babies invite ethical problems. The world began to CRISPR along and the bacterial immunity system becames a tool for genome editing by too many biotech startups.
-
The big lesson to aspiring 30 years young researchers in the names of Mojica, Horvath, Marraffini, Charpentier, Vogel, and Zhang working at not very famous places is the crux of the story. Journey from the repeat sequence curiosity to human genome editing is the climax of the CRISPR-CAS story. Big data bioinformatics helped. The lesson is that good discoveries are always the acts of an ensemble.
THE IMMUNE SYSTEM OF UNICELLULAR ORHANISMS
(7, 8,10, 18, 20, 23, 29,32, 34, 35, 37, 38, 41, 42, 43, 44, 45)
No doubt, CRISPR-Cas science has given a new tool for making and remaking of DNA. The story of CRISPR began with the finding of unusual repetitive sequences of the iap gene for alkaline phosphatase in E. coli (25). The repeat sequences later got their name as CRISPR (Clustered Regularly interspersed Short Palindromic Repeats). The repeats work with the DNA double strand breaking nuclease called Cas9 present in bacteria and halophilic archaea. The link between CRISPR and Cas9 enzyme system came about from the mobile genetic elements known as casposome or apoptosome that Cas9 knows how to bring about. This adaptive immunity system of bacteria has now become a tool for editing genes (6, 23). Cas9 is now being used to modify heterologous mammalian genes (56). It is a large gene with CRISPR and spacer which includes triplets to hexaplets of PAM (Protospacer Adjacent Motif). Cas9 breaks only DNA and not RNA (34). This large molecular weight enzyme has evolved by a team work with crRNA (complement of viral DNA as spacers) and tracrRNA (a separate RNA that can pair with crRNA) bound to Cas9 as one big complex (7). The enzyme alone has a volume of 500 cubic nanometers with N terminus recognition lobe and C terminus nuclease lobe. What better molecular intelligence there could be! The guide RNA that rides on it and acts as a target finder adds to the complexity of the structure. It is amazing that such a molecular complexity is required to fight a phage of limited coding capacity that invades a bacterium, navigates its DNA, replicates its own kind, and finally kills the host bacterium after using their replication machinery.
Let us get a grasp of the CRISPR structure that baffled scientists for a few decades. The repeat sequences of CRISPR with constant length generally have a dyad symmetry to form a palindromic structure (shown by arrows). Two examples are shown by the first identified CRISPR from E. coli (bacteria) and H. mediterranei (archaea). The spacer regions also have a constant length but without sequence homology.
Polymerase enzyme can make a complement to the template RNA (from DNA). Each snippet of CRISPR RNA contains a copy of a repeat and a spacer from a CRISPR region of DNA. The crRNA interacts with the Cas9 protein and another kind of RNA, called “trans-activating crRNA” or tracrRNA, in order to help the bacteria prevent from viral attack by destroying the DNA of the virus. Let us define these unique RNAs.

https://horizondiscovery.com/en/gene-editing/gene-editing-reagents/products/edit-r-predesigned-synthetic-crrna?gclid=Cj0KCQiA-oqdBhDfARIsAO0TrGFW5euiYfeh_k0lAGM5l5FIzAvpAdO7xRBFRqm9NtUQDrquk5OmG-oaAgDqEALw_wc
crRNA: This RNA is 17-20 db long and complementary to the spacer and the repeat. Also, crRNA, as a transcript of spacers, is complementary to the target DNA of the virus. It binds to Cas 9 proteins (17).
TRANSACTIVATING CRISPR RNA (traceRNA): Emmanuel Charpentier first described this small trans encoded RNA necessary for recognition and cleavage of an invading viral DNA, tracrRNA base pairs with crRNA for making a duplex structure into the gRNA. Only the duplex fits on to the Cas structure. In particular, it pairs with repeat sequence. It is a kind of anti-repeat as to separate locations of Palindrome repeat. It complements repeats, expressed separately by a promoter (15).
sgRNA is a single guiding RNA. crRNA is the part of gRNA (sgRNA) that is complementary to target DNA while the trcrRNA is the second part of gRNA (sgRNA) which serves a binding handle/scaffold for the endonuclease enzyme Cas9.
PAM is a component of the invading virus and not a component of the host bacterial DNA. There are two many varieties of PAM. Only a 2-6 bp long PAM (Protospacer Adjascent Motif) finds DNA region for cleavage by Cas9. For sure, Cas9 needs PAM. Cas9 may be engineered to recognize PAM. That would make CRISPR more efficient (54).
When a virus is nearby to attack, a copy of its DNA is integrated into the CRISPR locus. crRNA is transcribed from the CRISPR locus. crRNA bound to Cas guides it to the invading virus. At the present gRNA (sgRNA) is made artificially; it doesn’t exist in nature. The diagram below shows how the Cas-9 structure is configured to accommodate crRNA and TracrRNA on its specific lobes. Cas-9 bound to crRNA and tracrRNA thus is a nuclease with power to recognize target DNA and conduct double strand break.

https://bioscope.ucdavis.edu/2019/06/12/the-magic-cut-on-target-dna-by-crispr-cas9/
The Cas9-crRNA-tracrRNA complex binds to foreign DNA containing PAM. The endonuclease enzyme binds and starts to unwind the double strand of the foreign viral DNA to induce duplex formation of crRNA and foreign DNA. It consists of two regions, called the REC (recognition) lobe and the NUC (nuclease) lobe. The REC lobe is responsible for nucleic acid recognition. The NUC lobe contains the HNH and RuvC nuclease domains and a C-terminal region containing a PAM-interacting (PI) domain. The HNH and the RuvC domains of nuclease cleave the DNA strand, forming a duplex with crRNA and the other DNA strand, respectively, so that double-strand break occurs in the target DNA (SOURCE: https://sites.tufts.edu/crispr/crispr-mechanism/).
THE CRISPR ASSOCIATED NUCLEASE SYSTEM (19, 29,31, 40, 48, 53, 56)
To reemphasize, Cas9 protein is the nuclease that cuts foreign viral DNA. The protein binds to crRNA and tracr RNA, which together guide Cas9 to a target site on the virus’s DNA strand where the protein make the cut. The target DNA that the Cas9 will cut through is complementary to a 20-nucleotide stretch of the crRNA. Using two separate regions or “domains” (HNH and Ruuc) on its structure, Cas9 cuts both strands of the DNA double helix, doing what is known as a “double-stranded break.” “Protospacer adjacent motifs,” or PAM, serve as a tag and sits adjacent to the target DNA sequence. If the Cas9 complex doesn’t see a PAM next to its target DNA sequence, it won’t cut. This is one possible reason why Cas9 doesn’t attack the CRISPR region in bacteria. I include here the crystal structure of Cas9 Nuclease (Phys.org,References 46, 47)

https://www.sciencedirect.com/science/article/pii/S1097276517301211
To direct Cas9 to snip a specific region of DNA, scientists can simply change the sequence of the crRNA, which binds to a complementary sequence in the target DNA. In the 2012 Science article, Martin Jinek and his colleagues further simplified the system by fusing crRNA and tracrRNA to create a single “guide RNA/”. Thus, genome editing requires only two components: a guide sgRNA and the Cas9 protein.
BIOINFORMATICS AND CRISPR-Cas9 SYSTEM
Nature is very mysterious at the ultramicroscopic level. The adaptive immunological defense mechanism that the bacterial and the Archaea employ uniquely has taught us about a new tool for editing human and plant genomes respectively for human therapeutics and agricultural food productivity. The tool has multiple applications. To this effect, said Charpentier and Doudna, engineering of universal programmable RNA-guided DNA endonucleases creates the potential to exploit the system for RNA-programmable genome editing. The CRISPR-Cas9 double DNA break is similar to damages by free-radicals. Our cells can recognize a damaged DNA. Beyond the double strand break by CRISPR-Cas-9 system, we are at the mercy of our cell’s capacity to do the routine repair works. But first, let us examine the relevant bioinformatics that sets up the complexities of the human genome editing.
Twenty three (23) Pairs of Chromosomes, 46 double stranded 3 feet long (twisted and compressed) DNA helix per Cell, 8 histone proteins scaffolds per DNA, beads of chromatin complex on a string, 3.1 billion bases, and 20000 to 25000 genes represent information in the forms of bonds, bond strength and bond energies, shapes, complexes, configurations, and the topology of target DNA, RNA, and endonuclease molecules. Only an integrated set of information from genomics, transcriptomics, proteomics, metabolomics, and epigenomics leads us to credible personalized medicine, health, and nutrition. Every mammalian cell has 0.3 pg DNA, 20 pg RNA (1%) that needs more dedicated and revealatary research, 500 pg protein (20%), and 500 pg polysacharides (2 microgram). This information as bio-intelligence affords passive immunity to bacteria against viruses and now may help us develop therapies, medicine, high nutrition foods by the fast and simple methods of CRISPR-Cas human genome editing. This post unravels the stories of double strand breaking of a DNA molecules by systems of nucleases and RNA molecules that began with prokaryotes. It is now the dominant story of willful modification of human genome. Amazingly, this noble prize winning story that evolved in less than two decades is still ongoing. By all accounts the story is incomplete in terms of gene regulation and genomic markers. CRISPER-Cas system is enormously complex and evolutionarily mobile.
(https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5901762/) and
(https://www.sciencedirect.com/science/article/pii/S1672022914000278#:~:text=As%20one%20of%20the%20four%20major%20macromolecules%20(percentage%20weight%20in,roles%20in%20cellular%20processes%20and.)
A virus is 200 nm, a bacterium is 3000 nm, and our eukaryote cell is 30,000 nm (1:15:150 dimensional ratio).
The nucleus in the eukaryotic cell where chromosomes reside is 10,000 nm.
crRNA of CRISPR-Cas system is at the most a small, only 100 bases long (30 nm), roughly a fifth of the diameter of Cas9 enzyme. The length of tracrRNA is also small. Please the comparisons under A, B, and C below.
The molecular weight of Type II Cas9 endonuclease is 162 Kilodaltons (a volume of 10 nm × 10 nm × 5 nm from crystal structure) and a core diameter of 145nm (Chem. Sci. , 8(2): 7923-7931, 2017). Its rival protein is significantly different Class II type V CRISPR-Cas 12 a with its own pre-crRNA.
These two Cas systems have similar molecular weights, similar multidomain effectors of two lobe structure. Cas12a with only one nuclease site in the RuvC domain is reported to be more reliable than Cas9 with two domains but a complete repertoire of CRISPER-Cas system for a reliable human genome editing, it appears, has yet to evolve.

https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7090318/
C

https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7090318/
The largest human chromosome 1 is 248,956,422 base pairs or 10,000 nm long.
Human genes could be from 100 to a few million bases.
A bacterial gene is 0.6 to 0.8 million bases long capable of encoding 6000 proteins.
The bacterial genome is 0.1% of human genome size.
The bacteria have only 2500 genes compared to humans with 25,000 genes,
Now think of the intelligent logistics of a 500 nano cube Endonuclease Cas9 specific to a given bacterium with a bound 30 nm long crRNA traveling around more than 100 nm to the target DNA site to do a precise breaking of the two strands of DNA. We can visualize the scope of this scale in a picture from near atomic resolution cryo-electron microscope (Frederick National Lab for Cancer research). Such systems have been investigated in case of Yersinia pestris (50), Streptococcus thermophilus (51,53), and Heleobactor pylori (55). Plasmids (42) and Archaea (43) have been investigates as bacterial invaders also.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3497052/
Palindromes of the CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) are the key (2,3,6,9). The CRISPR-Cas9 uses the noncoding palindrome for recognizing the target DNA. gRNA guides the Cas9 endonuclease system to the site where the double strands of DNA needs to be broken prior to inserting new nucleotide snippets. Let us examine a more detailed explanatory diagram.

https://www.cell.com/molecular-therapy-family/nucleic-acids/fulltext/S2162-2531%2817%2930058-6
The CRISPR-Cas9 system: The sgRNA (purple) targets the Cas9 protein to genomic sites containing sequences complementary to the 5′ end of the sgRNA. The target DNA sequence needs to be followed by a proto-spacer adjacent motif (PAM), typically NGG. Cas9 is a DNA endonuclease with two active domains (red triangles) cleaving each of the two DNA strands three nucleotides upstream of the PAM. The five nucleotides upstream of the PAM are defined as the seed region for target recognition.
Spacers are transcribed into 17 to 20 bp long crRNA. The gRNA of the CRISPR-Cas9 system includes both the crRNA complementary to target DNA and tracr RNA which binds to Cas9 nuclease. The 24-28 bases long repeats separated by spacers of similar length are there to be used as Palindrome sequences for recognition.
A 20 bases long (can be synthetic) gRNA, based in a longer RNA scaffold, has bases complementary to target DNA and binds to it. Cas9 with the help of gRNA finds viral DNA that needs to be chopped. It is now well known that the number of the defense spacers correspond to increasing immunity against viruses. The virus doesn’t attacks bacteria that have genomes containing segments of snippets homologous to viral genome.
There are many CRISPR-Cas systems. The system’s action is usually divided into three stages: (1) adaptation or spacer integration, spacer being the segment of a foreign viral DNA, (2) processing of the primary transcript of the CRISPR locus (pre-crRNA) and maturation of the crRNA which includes the spacer and variable regions corresponding to 5′ and 3′ fragments of CRISPR repeats, and (3) DNA (or RNA) interference.
The invading DNA is recognized by Cas proteins during adaptation, fragmented and incorporated into the spacer region of CRISPR, and stored in the bacterial genome.
Pre-crRNA is generated by transcription of the CRISPR region and is processed and expressed into smaller units of RNA, named crRNA.
By taking advantage of the homology of the spacer sequence present in crRNA, foreign DNA is captured, and a complexed
with Cas protein nuclease that interferes with target DNA and cleaves it.
CRISPR-Cas genetic system is both complex and evolutionarily fast changing (Methods Molecular Biol., 1311: 47-75, 2015). There can be multiple CRISPR-Cas systems in the same organism. For instance, Streptococcus thermophilus has been found to have 4 different crRNA modules (Mol Microbiology, 93(1): 98-112, 2014). For now, we have Cas3 in type I systems, Cas9 and Cas 12 12 a2 in type II, and Cas10 in type III. Cas 12a2 is evolving fast. The three main types are readily distinguishable by virtue of the presence of three unique signature genes. The mechanism of CRISPR/Cas-9 genome editing contains three steps of recognition, cleavage, and repair. The designed sgRNA recognizes the target sequence in the gene (DNA) of interest through a complementary base pair.
The “information Processing modules” of Cas1 and Cas2 nucleases are present in the great majority of the known CRISPR-Cas systems. Either is sufficient for the insertion of spacers into the CRISPR cassettes, the a set of variable spacers regularly separated by palindromic repeats. CRISPR- Cas systems are quasi-autonomous by their complex molecular design that is required for this adaptation process. The endonuclease activity of Cas1 is required for spacer integration whereas Cas2 appears to perform a nonenzymatic function. Class 1 effectors contain multiple subunits while class 2 effectors are single large proteins. Cas3 like Cas9 is kind of a motor that finds the specific DNA target, runs along the offending DNA and conducts double strand break of DNA. Unlike Cas9, when Cas3 binds to its precise DNA target, it begins chewing up one strand of the double-stranded DNA in both directions, leaving a single strand exposed. Class 3 obviously is not ideal for engineering human therapeutics.
Cas9 requires only a “GG” sequence adjacent to its target. Cas9 can break the double strands of DNA at a site 3 base pairs upstream to PAM (protospacer adjacent to the motif) at a specific location where bits of DNA snippets can be added. The designed sgRNA recognizes the target sequence in the gene of interest through a complementary base pair. The double strand break is repaired by either non-homologous end joining or homology-directed repair cellular mechanisms.
The crRNA can be custom made. sgRNA (single guide RNA), is a single RNA molecule that contains both the custom-designed short crRNA sequence fused to the scaffold tracrRNA sequence. sgRNA can be synthetically generated or made in vitro or in vivo from a DNA template. Whereas crRNAs and tracrRNAs exist as two separate RNA molecules in nature, customized sgRNAs is now more commonly used. There may be reason to use cr:tracrRNAs (pronounced CRISPR tracer RNAs) for a specific objective. There are many software available for this purpose such as Synthego Design, Broad Institute GPP sgRNA designer, Crispor, CHOPCHOP, and CRISPR Era.
Just to restate, gRNA is the term that describes all CRISPR guide RNA formats, and sgRNA refers to the simpler alternative that combines both the crRNA and tracrRNA elements into a single RNA molecule. The diagram below shows tracrRNA binding to Cas9, crRNA complementing the target DNA, double strand cleavage of the taget DNA. The result is a finished new DNA.

https://www.researchgate.net/figure/Method-of-CRISPR-Cas9-action-and-epigenetic-manipulation-based-on-the-probability-to_fig4_342795533
The CRISPR-Cas9 System shown above is a ribonucleoprotein complex containing gRNA and Cas9 nuclease. A specific protospacer (white) adjacent motif (PAM) in the genomic (viral or human) DNA is required for the gRNA to bind to the target sequence. The Cas9 nuclease then makes a double-strand break in the DNA by scissor-k-like cut. Endogenous cellular repair mechanisms takes over thereafter. The figure below explains the editing process.

NEW DNA New DNA
https://www.addgene.org/crispr/cut/
The principle of genome editing is the cleavage of double-stranded DNA at a targeted position on a selected gene (DNA). For now the type II Cas-9 nuclease is the simplest of all nuclease-complexes among the CRISPR-Cas systems. Class II Cas 12a2 is evolving fast. The CRISPR RNA (crRNA), having a sequence homologous to the target site, and trans-activating CRISPR RNA (tracrRNA) are enough to bring the Cas9 nuclease to the target site. The artificial linkage of crRNA and tracrRNA (a complement to repeats) into one RNA chain (single-guide RNA [sgRNA]) has no effect on function. Once the Cas9-sgRNA complex cleaves the target gene, it is easy to disrupt the function of the gene by a deletion or insertion mutation. This simple genomic editing technique is found practical by most laboratories.
REENGINEERING GENES AND GENOMES (11, 14, 16, 17, 24, 28, 30, 33, 49, 52, 57, 59, 62, 65 )
Let us review the diagram below for reengineering details. CRISPR plasmids (vectors) or a CRISPER virus can be inserted into cell cytoplasm and can be transcribed in the nucleus. Also, Synthetic gRNA and Cas9 mRNA can be delivered to the cytoplasm. Cas-9 mRNA tranaslates into Cas-9 protein and the protein complexes with gRNA to make RNP complex which can enter the nucleus and do the DNA break. Alternatively, the entire Cas-9 and gRNA complex can be delivered to the cell cytoplasm and translocated into the nucleus.

https://en.vectorbuilder.com/products-services/service/crispr-offerings.html?gclid=CjwKCAiAzKqdBhAnEiwAePEjkl2xv3t8O9ickm5TBEYP98YmcvflllU5u5xfzuj6Qckulv4qbzjhwRoCeM0QAvD_BwE
CRISPR-Cas9, the bacterial immune system, can be created in a programable manner. A DNA-cutting enzyme (Cas9) and a programmable RNA molecule (guide RNA), call it CRISPR-Cas9, can be used to precisely target nearly any gene. Introduce the Cas9-guide RNA complex into a human cell where it randomly associates and dissociates with the DNA. Cas9 recognizes and binds to a three-nucleotide sequence motif called PAM that is abundant throughout the genome. One way to think of the Cas9-guide RNA complex is as a molecular scissor (Cas9) with a programmable GPS (guide RNA). Cas9 is a nuclease, a type of enzyme that cleaves DNA.
First we have to synthesize the guide RNA to contain a 20-nucleotide sequence that matches a particular sequence in a cell’s target DNA. When the guide RNA is added to Cas9, it will complex with and guide Cas9 to this target sequence. The target sequence can be nearly any sequence as long as it occurs near a PAM motif. it can be part of a gene’s coding region or a regulatory sequence that scientists want to change in some way.
The genetic material of an organism (DNA) is stored in the nucleus of cells. It consists of long sequences of nucleotides.
Guide RNA for a gene can be created in vitro. The motif consists of any nucleotide (designated “N”) followed by two guanines, when looking at a DNA sequence in a 5 prime to 3 prime direction (N-G-G). How often are the PAM sequences presented in the mammalian genome on average is a good question. Using the frequency of ‘GG’0f 5.21% in the reference human genome [1] there would be an expected 161,284,793 NGG PAM sites in the human genome, or roughly one ‘GG’ dinucleotide every 42 bases.
Once it binds to a PAM motif, Cas9 unwinds the DNA double helix. If the DNA at that location perfectly matches a 20-nucleotide sequence within the guide RNA, the DNA and matching RNA will bind through complementary base pairing. Cas9 recognizes and binds to PAM motifs in the cell’s DNA. After binding, Cas9 unwinds and pulls apart the DNA double helix upstream of PAM, closer to the 5 prime end of the DNA strand relative to PAM. If the sequence of the unpaired DNA strand is not an exact match to the 20-nucleotide sequence within the guide RNA, Cas9 disengages from the DNA, which zips back up into a double helix. If the sequences are a perfect match, the guide RNA base pairs with the complementary DNA sequence, forming a DNA-RNA helix.
Cleaving The DNA-RNA pairing triggers Cas9 to change its three-dimensional structure and activates its nuclease activity. When the guide RNA perfectly aligns with the target DNA, the RNA and DNA will form a DNA-RNA helix. This binding event activates Cas9’s nuclease for the DNA-cutting activity. Two active sites (regions where molecules bind to undergo chemical reactions) on the nuclease domain of Cas9 generate the cuts and cleave both strands of the DNA double helix, resulting in a double-stranded DNA break.
Our Cells contain enzymes that repair double-stranded DNA breaks. The repair process is naturally error-prone and will lead to mutations that may inactivate a gene. Therefore, cleaving DNA at a precise location is one of many applications of the CRISPR-Cas9 technology. Mutations or genome rearrangements occur if the DNA strands after double strand cut are not rejoined correctly. The sequence is very similar to the original sequence but the repair process can result in mutations, a change in the DNA sequence.
Gluing the two ends together (Huntington’s Outreach Project at Stanford University) is one repair method. This is known as “non-homologous end joining.” The problem with this method is that it tends to introduce errors where nucleotides are accidentally inserted or deleted, resulting in mutations that disrupt a gene. The break is fixed by filling in the gap with a sequence of nucleotides in the second method where the cell uses a short strand of DNA as a template. Scientists can supply the DNA template of their choosing, thereby writing-in any gene they want, or correcting any mutation of choice (Yoshizumi Ishino, Mart Krupovic, and Patrick Forterre,*J Bacteriol. 200(7), 2016). Non-homologous and homologous directed repairs are detailed further in the following diagram.

https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8444435/
NON-HOMOLOGOUS END-JOINING: CRISPR-induced double-stranded DNA breaks can be repaired by non-homologous end-joining (NHEJ). NHEJ is the more frequently used because it is faster repair mechanism. The cell does not use a template to join broken DNA ends together. It is, however, an error-prone process that can introduce mutations in the target sequence. Such errors are rare, but when the break is repaired correctly, Cas9 will once again recognize the target sequence and cleave it. Repeated cycles of cleavage and repair eventually result in a random mutation precisely within the target sequence. If the target sequence is within a gene’s coding region, the mutation will likely inactivate that gene. Mutations that inactivate a gene typically turn off its expression so that no protein is produced resulting in a nonfunctioning protein.
HOMOLOGY-DIRECTED REPAIR: The second type of repair mechanism is homology-directed repair (HDR), which is less error-prone and uses a homologous DNA template to accurately repair the break (for example, from a sister chromatid). Scientists can manipulate this repair system by introducing into the cell an excess of a DNA repair template along with the Cas9- guide RNA complex. The cell’s repair machinery will be “tricked” into using the repair template to fix the break by HDR. By designing different repair templates, scientists can change the target DNA sequence into a new sequence. These templates could also correct an existing mutation by replacing it with a non-mutated sequence of DNA. DNA sequences can be inserted, modified, or deleted. A gene can be added or deleted, a small section of DNA can be changed, and even gene expression can be refined. Scientists have successfully edited single cell disease mutations.
Human Transcriptome (a DNA to mRNA Map) generates gene expression profiles for any chromosomal region in 12 normal and pathologic tissue types. The map reveals a clustering of highly expressed genes to specific chromosomal regions. It provides a tool to search for genes that are overexpressed or silenced. The human transcriptome map reveals extremes in gene density, intron length, GC content, and
repeat pattern for domains of highly and weakly expressed genes (https://genome.cshlp.org/content/13/9/1998.full.pdf).
Also, the genomes encode a series of messages and instructions within their DNA sequences and genome editing involves changing those sequences, thereby changing the messages they contain. This can be done by inserting a cut or break in the DNA and tricking a cell’s natural DNA repair mechanisms into introducing the targeted changes. CRISPR-Cas9 system offers a simple means to do so. “Operationally”, one designs a stretch of 20 base pairs that match a gene that you want to edit and, from there, one can figure out what the complementary crRNA-like sequence would be. The nucleotide sequence of choice is found only in the target gene and nowhere else in the genome.
CUUREENT STATE OF REPURPOSED TECHNOLOGY
I like the diagram below. It details the name of establishments along with names of scientists in the outermost perifery of the circle followed by centers of research and intellectual property development, and finally the areas of applications.

labiotech.eu/ in-depth/crispr-cas-review-gene-editing-tool/
Crisper Therapeutics, founded by Emmanuel Charpentier, operates both in USA and Switzerland. It works with Virtex Pharmaceuticals trying to solve the blood disorder problems. This company’s approach is EX VIVO technology of extracting the relevant cell from a patient, reengineering it in the lab and finally introducing back in the patient’s body. The IN VIVO technology that Intellia Therapeutics , founded by Janiffer Doudna, uses is directed to neurological diseases like transthyrectin amyloidosis. Editas, founded by Feng Zhang, is working on blindness and cancer. There are many delivery methods uses today as listed in the table below.

https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8444435/
CRISPR-Cas delivery methods.
Major categories |
Delivery vehicle |
Advantages |
Limitations |
Physical delivery methods |
Microinjection |
Guaranteed delivery into cell |
Difficult to operate |
Electroporation |
Delivery to cell population |
Generally in vitro only |
|
Hydrodynamic delivery |
Low cost |
Traumatic to tissues |
|
Membrane deformation |
Suitable for a variety of target cells |
High cost |
|
Viral vectors |
AAV |
Minimal immunogenicity |
Low capacity |
AdV |
High efficiency delivery |
Inflammatory response |
|
LV |
Persistent gene transfer |
Prone to gene rearrangement |
|
Nonviral vectors |
Nanoparticles |
Virus-free, reformable |
Require complex design and more researches |
Cationic lipids |
Virus-free, simple manipulation |
Toxicity |
|
Exosomes |
High biocompatibility, low immunogenicity |
Low loading efficiency |
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8444435/
The RNA guided Class II Cas-9 is 1368 amino acid long with MW of 160 KD. This is the first nuclease protein isolated from Streptococcus pyogenes. Formerly, also called Cas5, Cas 13, Csn1, and Csx12/, the CRISPR-Cas9, is an important immunological defense of bacteria against viruses/phages and plasmids. The signature gene for type II CRISPR-Cas 9 systems encodes a multidomain protein that combines all the functions of an effector. Adapted from the natural immunity mechanism of single cell bacteria, it can now be used for restructuring the genes of eukaryotic genomes. The current emphasis is on human therapeutics and cell engineering.
Both RNAs can be synthesized. The crRNA is an 18–20 base pair in length that specifies the target DNA by pairing with the target sequence and tracrRNA is a long stretch of loops that serve as a binding scaffold for Cas-9 nuclease.
In case of prokaryotes, the guide RNA is used to target viral DNA, but as a general gene-editing tool, it can be synthetically annealed as a single guide RNA in order to target almost any gene sequence that we need to edit.
The molecular mechanism of the target DNA cleavage by the Cas9-crRNA complex has been elucidated at the near atomic level by the crystal structure analysis of the DNA-Cas9-crRNA complex. This genome-editing tool is a huge discovery of 2012 with wide number of applications in many areas including agriculture, medicine, food fermentations, and biotechnology. In agriculture, it could help in the design of new grains to improve their nutritional value. There is talk of creating a caffien free coffee plant. In medicine, it is being investigated for treatment of cancers, HIV, and gene therapies for sickle cell disease, cystic fibrosis, and Duchenne muscular dystrophy by modifying the human genome. The CRISPR-Cas-9 template can change easily and speedily any gene anywhere. Same Cas-9 with different synthetic RNA does the work. Although CRISPR/Cas-9 stands out as a new era in molecular biology with countless roles ranging from basic molecular researches to clinical applications, it faces challenges to cope up with (biologics, 15: 353–361, 2021). Class II Cas 12 a2 is now being research as an alternative tool.
The genetic renaissance can be summed up as training a yeast cell to produce almost complete synthetic chromosome, producing pluripotent stem cells without codes for individuality, parsonalysing to a specific patient, and have T-cells fight a given cancer.
There are close to 1221 clinical trials, most of them in stages 1 and 2, on disease treatments by CRISPR gene editing. Target area are blood disorders of sickle cell anemia and beta thalassemia, hemophilia, cancers, inherited eye disease, Type 1 Diabetes, Infectious diseases, inflammatory diseases, and protein folding disorders. Immune cells can be altered such that they recognize tumor specific proteins,
Dr. Robert Chiesa of Great Ormond Hospital in London has treated aggressive leukemia by CRISPER edited T-Cells. The patient is a 13 year old girl and she is in remission . Crisper Therapeutics has filed a trial with FDA on Type 1 Diabetes. There are three CRISPER babies pseudonamed Amy and twins Lulu and Nana in China by Dr. Jiankui who was imprisoned on grounds of unethical research.
PRACTICAL APPLICATIONS
CRISPR-Cas has its beginning with yogurt bacteria in the hands of Phillipe Howarth and Rodolphe Barrangou at Danisco in France. Also, CRISPER-Cas9 tool has been used to improve yeast strains for wine making and Japanese scientists are busy perfecting Aspergillus oryzae for better soya sauce.
Yogurt by lactic bacteria is a 7000 years old fermentation invented, like butter, by accident. India’s Dahi is even older. Origin of wine fermentation by yeast goes back 6000 BC in S Caucus, Georgia. Beer’s origin by yeast fermentation goes back 4000 BC in Mesopotamia in the city of Babylon. Bread by yeast fermentation began in Egypt around 1000 BC. Blue cheese fermentation by bacteria and fungus is a 7th century accidental invention in the caves outside Roquefort in France. The Jan of China and Hisio of Japan got perfected from miso paste to soy sause by 1688. Sour Kraut by lactic fermentation (old sour cabbage) was perfected as a fermented food by World War I. Penicillin was invented by Alexander Fleming at the St Mary’s Hospital in London rather recently in 1928. In theory, all these fermentations can be improved by the CRISPR technology and CRISPR-Cas9 system is already on the minds of food scientists all over the world. Improved food fermentations may provide for more nutritious foods for the expanding population of the world. Let us not forget that viruses are also our friend. We use Retro-viruses and Lentiviruses for the delivery of CRISPER-Cas9 load to human nucleus.
I learned about the devastating infections by these phages in butter and cheese plant in 1963. My 55 years old intimacy with phages tells me that while they can be very destructive to a $157.00 billion global industry of butter, cheese, and vinegar (respectively $ $35.61, $120.63, and $1.32 respectively), they can potentially rescue the industries of $1.34 trillion beer, wine, and soy sauce industries (respectively $680.00, $456.00, and $47.00 billion) by preventing Lactobacilus plantarum like contaminations. Economically, this benefit seems to scope out better than the urgently needed use of CRISPR-Cas9 system for therapeutics and prevention of human diseases. Most investments are directed to the latter. Let me include a list of known applications. Our readers may like to consult (Precis Clin Med. 2021 Sep; 4(3): 179–191) and URL
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8444435/ beyond the list.
-
General and Germ Line Gene Editing
-
Designer babies
-
Cancer Immunotherapy
-
Heart disease cure
-
Cholesterol reduction
-
Prevent fungal attack on chocolate production
-
Diagnostic tests for COVID-19 pandemic and Zika
-
Prevent diseases of Huntigton’s and sickle cell anemia
-
New more powerful antibiotic production
-
Gene Drives of Wyss Institute for wiping out malaria mosquitoes.
-
Once in life time vaccine against chronic diseases
-
Pet Breeding
-
New herbicide resistant crops
-
Poultry Breeding for better Foods (https://www.frontiersin.org/articles/10.3389/fgene.2021.627714/full)
-
Allergy Free Foods.
-
Production of tasty, nutritious, and and flavorful foods
-
Improved food fermentations: yogurt, cheese, soy sauce, and sour kraut production.
-
Spicy Tomatoes
-
More nutritious salmon
-
Alligator genes in fish prevent fish diseases.
-
Greener Fuel/ Bioenergy
-
Faster Race Horses and Pet Breeding
-
De-Extinction
-
DNA Tape Recorders (https://singularityhub.com/2018/02/27/not-just-gene-editing-crispr-toolkit-expands-with-trio-of-new-tricks/#sm.001tr8wct1blbdztu7616o2ctf15t). This amounts to transforming the hotshot editor into a virus detective or a sharp-eyed historian jotting down a cell’s entire history into DNA.
Here are the steps to gene editing for therapeutics or developing new organisms or crops.
Steps in CRISPR-CAS9:
-
Select an organism for the experiment.
-
Select a gene of the target location.
-
Select a CRISPR-CAS9 system.
-
Select and Design the sgRNA.
-
Synthesizing and cloning of sgRNA.
-
Delivering the sgRNA and CAS9.
-
Validating the experiment.
-
Culturing the altred cells.
THE FUTURE OF CRISPR-Cas9 GENETICS
All that lives evolves with genes and gene editing with CRISPER-CAS systems could potentially change all. The structure of CRISPR-Cas below is complex. We do not know all of it’s functional ramifications. It may not always fire the way we want it to and there in lies the danger. But then there are two many types to cut DNA and RNA strands from which to select the ideal one for a given application.

https://www.aamc.org/news-insights/future-crispr-now
The first EX VIVO trial for cancer treatment began in 2018 and immediately thereafter in 2019 began the a IN VITRO trial was approved by FDA. The simplicity of CRISPR-Cas9 notwithstanding, it is going to take some time before it is used routinely for human therapeutics. Research is also focusing on its use in animal models or isolated human cells, with the aim to eventually use the technology for routine treatment of human diseases. Expressions may not always be specific as to gene correction though. A lot of work is focused on eliminating ‘off-target’ effects that result from CRISPR-Cas-9 system cutting at a different gene and not the one that was intended to be edited. Once we find an ideal gRNA, CRISPR-Cas-9 system will do a great job.. This may help evolve standard clinical care methods.
CRISPR-Cas9 must bind and cut accurately by more specific guide RNAs based on our knowledge of the DNA (gene) sequence and the ‘off-target’ behavior of different versions of the Cas9-gRNA complex must be rigorously controlled. Current focus is on genotyping, vaccination, controlling uptake, and engineering the CRISPR-Cas systems.
In most cases the guide RNA consists of a specific sequence of 20 bases. These are complementary to the target sequence in the gene to be edited. However, not all 20 bases need to match for the guide RNA to be able to bind. A sequence with, for example, 19 of the 20 complementary bases may exist as a gene somewhere at a completely different location in the genome. This means there is potential for the guide RNA to bind there instead of or as well as at the target sequence and cut at the wrong site and end up introducing a mutation in the wrong location. While this mutation may not matter at all to the individual, it could affect a crucial gene or another important part of the genome because many genes work together.
The use of a Cas9 enzyme that will only cut a single strand of the target DNA rather than the double strand poses may open extra-opportunities. It would require two Cas9 enzymes and two guide RNAs to be present in the same place for the cut to be made. The technique will reduces the probability of the cut being made in the wrong place.
Instability of Cas9-DNA complex, specially when other motor proteins like RNA polymerase and translocases are also running around in the vicinity. Furthermore, the R-loop on the diastal side of PAM that is critical to binding may have to be strengthened (Peter M Hall etl.m Nature Structural Molecular Biology, December 5, 2022). I believe that most of these problems will be solved soon and a successful gene therapeutics will evolve that is low cost, simple, specific, portable, easy to operate, safe, and effective. Nobel Laureates Jennifer Doudna and Emmanuel Charpentier have founded their startups with exactly that in mind.
APPENDIX I: CLASSIFICATION OF CRISPR-Cas SYSTENS (36)

https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8444435/
First of all we do not have a rational nomenclature for the CRISPR-Cas systems. The systems are complex subject to fast evolution, modular suffling, and lack of universal markers. They are diverse in gene composition and the genomic locus architecture and even the sequences. Often he repeats show show clustering. There are no universal Cas genes and the current classification is based on signature genes specific to a type and subtypes. Class 1 is most conserved. The current classification goes by effector genes with numerous accessory genes genes with secondary roles. The modular organization changes without loss of functions. The 13 groups whose adaptation and effector modules we know represent a minority. The current classification will stay in place as long as we do not have an understanding of their structures (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6636873/).
The immunity system of CRISPR loci can change within a month. Bacteria and plasmids use CRISPR-derived RNA, complementary to its DNA, and various Cas proteins to fight attacks by viruses. CRISPR array in a given genome may have 1 to 18 repeats interspersed with spacers that are the viral DNA homologues. It is CRISPR that is transcribed into g RNA. A bacterium chops up the DNA of viruses and then stows bits of that DNA in their own genome, to be used as a weapon against new viral invaders in future. Different Cas enzymes have different activities. A convenient classification of important Cas types is outlined below.
CLASS I- TYPE I: 7 subtypes, 4 to 7 multi-unit Cas3 endonuclease that cleaves SS DNA strands. It doesn’t require Tracer RNA and its target is DNA only. Presnt in E. coli.
CLASS I -TYPE III: has 4 subtypes, this Cas10 multi-unit endonuclease binds to a nascent RNA, doesn’t require Tracer-RNA, and targets both RNA and DNA. IIIB is present in P. furiosus and IIIA is present in S, epidermidis.
CLASS I-TYPE IV ( Least Understood): has one subtype and its CTISPER association, targets, and Tracer-RNA requirements are not known.
CLASS II-TYPE II: This Cas9 endonuclease has 3 subtypes, is single unit, originates blunt DSB, targets DNA only, does require Tracer-RNA, and is present in 10% bacterial loci. Cas9 is most widely used today. Present in streptococcus thermophilus.
CLASS II- TYPE V: This Cas12 single multidomain unit endonuclease of bacteria has 3 subtypes, originates staggered DSB, targets DNA, and does require Tracer-RNA. Present in F. novicida’s cpf1 system.
CLASS II-TYPE VI: This single unit Cas13 endonuclease is an RNA guided RNase, targets RNA, and doesn’t require Tracer-RNA. SOURCE: https://www.synthego.com/blog/crispr-systems and https://www.ibl-america.com/blog/the-era-of-crisprcas-technology/
Type II is the simplest of all and Cas 9 is a single multidomain protein guided by the duo of CrRNA and tracrRNA for double strand break of the target DNA. It has six domains: RECI, REC I, Bridge helix, PAM interfering HNH, and RuvC.

site.tufts.edu/crispr/crispr.mechanism/
APPENDIX II: CRISPR-Cas VOCABULARY
Allele: One of several possible versions of genes with variance in DNA sequence.
Allogenic: Cell from different individuals of same species.
Archaea: bacteria like unicellular organism that are closer to Eukaryotes.
Autologous: Cells from same individuals.
Autosome, Largest is chromosome 1, gene on the same numbered non-sex chromosome.
BLUNT DSB: Post double stranded break, both strands are of equal length without any unpaired base.
Cas: CRISPR Associated. Cas from different organisms recognize different PAMS. Cas protein recognizes the invading DNA, frgments it, and incorporates into the CRISPR locus.
Casposome: The multiprotein complex casposome (apop- tosome) is comprised of cytochrome C, Apaf-1, pro- caspase-9 and ATP
Centromere appears as a constricted region of a chromosome and plays a key role in helping the cell divide up its DNA during division (mitosis and meiosis).
Codons: nucleotide sequence that codes aminoacids. The coding process proceeds from 5′ to 3′ end.
Conserved Sequence: A base sequence in a DNA molecule (or an amino acid sequence in a protein) that has remained relatively unchanged throughout evolution.
Chromatid is one of the two identical halves of a chromosome.
Chromatin refers to a mixture of DNA and proteins that form the chromosomes found in the cells of humans and other higher organisms.
CRISPR (24-28 nt long Clustered Regularly Interspersed Short Palindromic Repeat) in bacteria or archaea is a region of DNA made of short repeat sequences (ordering of base pairs) with spacers sandwiched between the repeats. Adenine and Thymine is a rung and so is Cytosine and guanine. Repeat sequences which read the same forward or backward have been called “palindromic” by the Max Planck Institute. The spacers are segments of virus DNA that serve as memory bank. This helps bacteria to recognize the invading virus. The noncoding palindrome gene is the key in CRISPR functionality. CRISPR and Cas genes work together.
• Side 1 – GATC
• Side 2 – CTAG
CrRNA: CRISPR RNA, also known as gRNA
Dicer is a dsRNA ribonuclease similar to bacterial Class III RNase III that comprises an N-terminal ATP-dependent RNA helicase domain, a PAZ (piwiargonaute-zwille) motif, and two C-terminal endonuclease domains followed by a single dsRNA binding domain.
DOUBLE STRAND BREAK: Cleavage of the phosphodiester link that often requires many enzymes. The shape of strands at the terminus matters. Double strand breaks can be detected in just a few hours by immunofluorescence staining of the phosphorylated histone H2AX. Such breaks are most cytotoxic.
Dyad Symmetry: two areas of DNA Strands whose base pair sequences are inverted repeats of each other.

Sciencedirect.com
Epistasis is a circumstance where the expression of one gene is modified (masked, inhibited or suppressed) by the expression of one or more other genes.
Eukaryotes: Multicellular organisms with nucleus in their cells.
Exome is the sequence of all the exons in a genome, reflecting the protein-coding portion of a genome. In humans, the exome is about 1.5% of the genome. Exon is a region of the genome that ends up within an mRNA molecule. Some exons are coding, in that they contain information for making a protein, whereas others are non-coding. Genes in the genome consist of exons and introns.
Expression can involve either transcription of DNA into RNA or translation by RNA into protein.
gRNA binds to target viral DNA.
Gamete is a reproductive cell of an animal or plant, e.g., egg and sperm.
Gene is considered the basic unit of inheritance. Genes are passed from parents to offspring and contain the information needed to specify physical and biological traits. Most genes code for specific proteins, or segments of proteins, which have differing functions within the body. Humans have approximately 21,000 protein-coding genes. The number has been various reported even up to 25,000.
Gene expression is the process by which the information encoded in a gene is used to either make mRNA molecules that code for proteins or to make non-coding RNA molecules that serve other functions. Gene expression acts as an “on/off switch” to control when and where RNA molecules and proteins are made and as a “volume control” to determine how much of those products are made.
Gene mapping refers to the process of determining the location of genes on chromosomes by complete sequence analysis.
Gene regulation is the process used to control the timing, location and amount in which genes are expressed.
Genomic marker or a genetic marker is a DNA sequence, typically with a known location in a genome.
Genome is the entire set of DNA instructions found in a cell. In humans, the genome consists of 23 pairs of chromosomes located in the cell’s nucleus, as well as a small chromosome in the cell’s mitochondria. A genome contains all the information needed for an individual to develop and function.

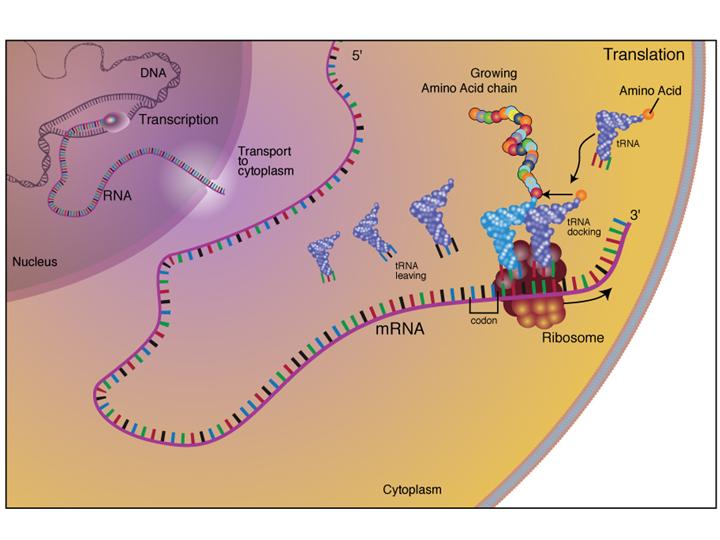{kind=link}
{kind=link}
https://www.istockphoto.com/vector/cell-chromosome-dna-and-gene-gm506992506-84490151?phrase=chromosome
Gene Therapy: CRISPR-Cas9 Therapy beyond those based on lentivirus or Adenovirus.
Guide RNA (gRNA): A two-piece molecule that Cas9 binds and uses to identify a complementary DNA sequence. It is a transcript of both the repeats and the spacers, composed of the CRISPR RNA (crRNA) and trans-activating CRISPR RNA (tracrRNA). Cas9 uses the tracrRNA portion of the guide as a handle, while the crRNA spacer sequence directs the complex to a matching DNA sequence. Scientists have also formed a version of the guide RNA that consists of a single molecule, the single-guide RNA (sgRNA).
HNH DOMAIN: A region of Cas9 known as the HNH domain is responsible for cutting the target DNA.
Haploid refers to the presence of a single set of chromosomes in an organism’s cells. Sexually reproducing organisms are diploid (having two sets of chromosomes, one from each parent). In humans, only the egg and sperm cells are haploid.
Helicases are defined as a class of enzymes that catalyze the separation of duplex nucleic acids into single strands in an ATP-dependent reaction and function in DNA modification processing, including DNA replication, DNA repair, recombination, transcription, translation, and many other nucleic acid-related processes. The enzymes break the hydrogen bonds and separate the strands.
Heterozygous refers to having inherited different versions (alleles, a mutated sequence) of a genomic marker from each biological parent.
Histone is a protein that provides structural support for a chromosome. Each chromosome contains a long molecule of DNA, which must fit into the cell nucleus. To do that, the DNA wraps around complexes of histone proteins, giving the chromosome a more compact shape. Histones also play a role in the regulation of gene expression.
Homologous recombination is a type of genetic recombination in which nucleotide sequences are exchanged between two similar or identical molecules of DNA. During the formation of egg and sperm cells (meiosis), paired chromosomes from the male and female parents align so that similar DNA sequences can cross over, or be exchanged, from one chromosome to the other. This exchanging of DNA is an important source of the genomic variation seen among offspring.
Homozygous refers to having inherited the same versions (alleles) of a genomic marker from each biological parent.
Homology-directed repair (HDR): A way for a cell to repair a break in its DNA by “patching” it with a piece of donor DNA. The donor DNA must contain similar sequences, or homology, to the broken DNA ends for it to be incorporated. HDR is a more precise repair pathway than non-homologous end joining. In genome engineering, a researcher designs and adds in the donor DNA, potentially allowing to replace a disease-causing gene with a healthy copy.
Hybridization is the process in which two complementary single-stranded DNA and/or RNA molecules bond together to form a double-stranded molecule. The bonding is dependent on the appropriate base-pairing across the two single-stranded molecules. Hybridization is an important process in creating new genes.
Intergenic regions are the stretches of DNA located between genes.
Intron is a region that resides within a gene but does not remain in the final mature mRNA molecule following transcription of that gene and does not code for amino acids that make up the protein encoded by that gene. Most protein-coding genes in the human genome consist of exons and introns.
Inversion in a chromosome occurs when a segment breaks off and reattaches within the same chromosome, but in reverse orientation. DNA may or may not be lost in the process.
Karyotype is an individual’s complete set of chromosomes.
Linkage refers to the closeness of genes or other DNA sequences on the same chromosome.
Locus is a physical site or location within a genome (such as a gene or another DNA segment of interest), somewhat like a street address.
Lyonization (named after British scientist Mary F, Lyon) refers to the normal phenomenon in which one of the two X chromosomes in every cell of a female individual is inactivated during embryonic development. This inactivation prevents females from having twice as many X chromosome gene products as males, who possess only a single copy of the X chromosome.
mRNA is at the the 5’ end capped with a guanosine triphosphate nucleotide for recognition purposes. Makes only 5% of total RNA in a cell.
Methylation is a chemical modification of DNA and other molecules that may be retained as cells divide to make more cells. When found in DNA, methylation can alter gene expression.
Major and Minor Grooves of DNA: Two distinct grooves known as the minor and the major grooves form on the opposite sides of the base pairs. The formation of minor grooves takes place where the sugar phosphate backbones are far apart. While the formation of major grooves takes place where the sugar phosphate backbones are close together. Very important for protein binding.

Microsatellite refers to a short non-coding segment of DNA, usually one to six or more base pairs in length, that is repeated multiple times in succession at a particular genomic location.
Nucleosome is the basic repeating subunit of chromatin packaged inside the cell’s nucleus. In humans, about six feet of DNA must be packaged into a nucleus with a diameter less than a human hair, and nucleosomes play a key role in that process. A single nucleosome consists of about 150 base pairs of DNA sequence wrapped around a core of histone proteins. In forming a chromosome, the nucleosomes repeatedly fold in on themselves to tighten and condense the packaged DNA.
PAM ( Protospacer Adjacent Motif ) are 3-4 bases long.
Plasmid is a small circular DNA molecule found in bacteria and protozoans. Plasmids are non-chromosomal DNA and replicate independently. Typically, they have a small number of genes — notably, some associated with antibiotic resistance. They can be passed from one cell to another. Plasmids are good gene carriers in DNA modification by recombination.
Restriction Enzyme is a protein isolated from bacteria that cleaves DNA sequences at sequence-specific sites, producing DNA fragments with a known sequence at each end. The use of restriction enzymes is critical to certain laboratory methods, including recombinant DNA technology and genetic engineering.
Retrovirus is a virus that uses RNA as its genomic material. Upon infection with a retrovirus, a cell converts the retroviral RNA into DNA, which in turn is inserted into the DNA of the host cell. The cell then produces more retroviruses in case of diseases like AIDS and some forms of cancer.
TYPES OF RNA : mRNA, the largest, Ribosomal RNA, tRNA, and SI RNA.
Ribosomal RNA (rRNA), molecule in cells that forms part of the protein-synthesizing organelle known as a ribosome and that is exported to the cytoplasm to help translate the information in messenger RNA (mRNA) into protein.
Ribosome is a 20-30 nm diameter complex structure made of both RNA and protein. It is the site of protein synthesis in the cell. The ribosome reads and rides the messenger RNA (mRNA) sequence and translates that genetic code into a specified sequence of amino acids which grow into long chains that fold to form specific proteins. Its 50s and 30s units are made of their own specific RNAs.
SEX Chromosome is a type of chromosome involved in sex determination. Mammalians including humans have two sex chromosomes, X and Y. Females have two X chromosomes in their cells, while males have one X and one Y.
Single-guide RNA (sgRNA): A version of the naturally occurring two-piece guide RNA complex engineered into a single, continuous sequence. The simplified single-guide RNA is used to direct the Cas9 protein to bind and cleave a particular DNA sequence for genome editing. sgRNA is custom made as a fused crRNA and tracrRNA.
siRNA is a non-coding double-stranded RNA molecule. It is also known as silencing RNA and short interfering RNA. It is similar to microRNA (miRNA) and the structure is short and well-defined, usually between 20 and 24 base pairs. Their structure has hydroxylated 3′ and phosphorylated 5′ ends.
Spacers are complementary to viral DNA, 17-19 nt long.
Telomere is a region of repetitive DNA sequences at the end of a chromosome. Telomeres protect the ends of chromosomes from becoming frayed or tangled. Each time a cell divides, the telomeres become slightly shorter. Eventually, they become so short that the cell can no longer divide successfully, and the cell dies. We need to keep them long for long life-span.
Transcription is the process of making an RNA copy of a gene’s DNA sequence. This copy, called messenger RNA (mRNA), carries the gene’s protein information encoded in DNA (sequence of nucleotide triplet to an amino acid). In humans and other complex organisms, mRNA moves from the cell nucleus to the cell cytoplasm so it is readily available for protein synthesis.
Trans-activating CRISPR RNA (tracrRNA): Abbreviation for trans-activating CRISPR RNA, pronounced “tracer RNA.” In the CRISPR-Cas9 system, the tracrRNA base pairs with the crRNA to form a functional guide RNA (gRNA). It binds to Cas9. Cas9 uses the tracrRNA portion of the guide as a handle, while the crRNA spacer sequence directs the complex to a matching viral sequence. tracrRNA has short hairpins.
Transfer RNA (tRNA) is a small 76 to 93 nt long RNA molecule that is the key to protein synthesis. It binds to the anticodon of mRNA. There are 20 t0 31 of them. It links the messenger RNA (mRNA) molecule with the growing chain of amino acids as protein. Each time an amino acid is added to the chain, an amino acid specific aminoacyl-tRNA pairs with its complementary sequence on the mRNA molecule. This ensures appropriate amino acid insertion in the amino acid sequence of the protein. tRNAs have a cloverleaf structure which is stabilized by strong hydrogen bonds between the nucleotides. They normally contain some unusual bases in addition to the usual 4, which are formed by methylation of the usual bases. Methyl guanine and methylcytosine are two examples of methylated bases.
Translation is the process by which information encoded in messenger RNA (mRNA) directs the sequential addition of amino acids during protein synthesis. It takes place on ribosomes in the cell cytoplasm, where mRNA is read and translated into proteins made of amino acids.
Translocation occurs when a chromosome breaks and the (typically two) fragmented pieces re-attach to different chromosomes. Translocations causes genetic diseases and disorders. AUG for methionine is the initiation codon.
RuvC DOMAIN: The RuvC Nuclease initiates cleavage of the DNA strand not complementary to the guide RNA
APPENDIX III: CHRONOLOGY AND TIME-LINE OF CRISPR R & D
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8036902/
More often than not seminal research comes about by budding young minds. In case of CRISPR, Mojica, Horvath, Marraffini, Charpentier, Vogel, and Zhang were early in their carrier below 30 years of age when their pursuit of this disruptive science began. Apparently so disruptive that their papers were initially rejected. Even in the absence of great hypotheses, big data helped them make their case and the new CRISPR science blossomed in less than 20 years leading to a Nobel Prize to Carpentier and Doudna. Feng Zhang of the Broad Institute received the First CRISPR patent. What evolved in a laboratory in Spain 20 years ago went Viral. Scientists found the immune system of CRISPR in fungi, nematodes, yeast, fruit fly, zebrafish, mouse, and monkey also. Currently there is a great emphasis on human therapeutics and Agricultural Productivity by methods of CRISPR-Cas gene editing.
The discovery of a strange microbial repeat sequence; its recognition as an adaptive immune system; its biological characterization; and its repurposing for human genome engineering has a been the dominant trend in CRISPR-Cas research.
Repurposed into a simple and reliable technique for editing the genomes of mammals and other organisms in vitro or in vivo is in focus and priority. CRISPR-cas9 system is now adapted for a vast range of applications for creating complex animal models of human-inherited diseases and cancers; performing genome-wide screens in human cells to pinpoint the genes underlying biological processes, turning specific genes on or off; and genetically modifying plants. Type II CRISPR-Cas-9 research is being done in thousands of labs worldwide.
Major workers on the CRISPR-Cas system include Barrangou and Marraffini (2014), Hsu et al. (2014), van der Oost et al. (2014), Sander and Joung (2014), Jiang and Marraffini (2015), Jiankui, Emmanuel Charpentier, Sternberg and Doudna (2015), Wright et al., 2016), and Fiang Zhang. A chronology of land mark research is listed below.
UNUSUAL REPEATS IN BACTERIAL DNA ARE NOW DEFINED AS CRISPR
1987: Amemura Ishino etal of Osaka University first report on clustered repeats. The “iap” gene in gut bacteria E. coli was sequenced to find that the DNA segment used was longer than the gene itself. They accidentally discovered a partial DNA sequence of then-unnamed CRISPR in the process, which eventually became the basis of CRISPR gene editing.
1993- Ishino Y. etal. Discovered CRISPR in Archeae Haloferax mediterranei.
1993 – 2005 — Francisco Mojica, University of Alicante, Spain, found the the CRISPR locus and noted that Spacers are snippets of virus DNA sequence and that CRISPR is an adaptive immune system. Pourcel et al., (2005) reported the same .
CRISPR FOUND TO BE PRESENT IN ALL EUKARYOTES
2000: Francisco Mojika declared that CRISPR is present throughout the prokaryotes.
CRISPR IS THE SIGNATURE GENE FOR Cas
2002: Jansen defined CRISPR as the signature CRISPR-Associated gene and identified CRISPR-Cas9.
2005: Mojika and Alexander Balotin of French National Institute for Agricultural Research (INRA) identified foreign origin of spacers.
Bolotin found in a Streptococcus thermophilus study the complete genome sequence with CRISPR LOCUS along with spacers homologous to viral DNA segments. All spacers shared a common sequence at the end, the protospacer adjacent motif (PAM) required for target recognition. This offered a hypothetical scheme of adaptive immunity. Soon the gene for endonuclease were discovered. The protospacer adjacent motif (or PAM for short) is a short DNA sequence (usually 2-6 base pairs in length) that follows the DNA region targeted for cleavage by the CRISPR system, such as CRISPR-Cas9.
SPACERS FOUND TO BE VIRAL DNA HOMOLOGUES
March, 2006 — Eugene Koonin at the US National Center for Biotechnology Information of NIH showed experimentally that spacers are homologous DNA fragments of virus for adaptive immunity.
CRISPEER IS AN ADAPTIVE IMMUNE SYSTEM
March, 2007 — Philippe Horvath at Danisco in France experimented with Steptococcus thermophilus in Yogurt and cheese and confirmed that CRISPR IS an adaptive immune system against next viral attack. It was shown that Cas9 is the only nuclease protein required. Rodolphe Barrangou and a team of Danisco Researchers and Department of Energy (2007) also worked on Streptococcus thermophilus of yogurt attacked by virus and found that there was additional new spacer identical to virus genome. When they removed the spacer and added new viral DNA, the bacterial immunity changed (22).
SPACERS AS VIRAL DNA ANALOGUES ARE TRANSCRIBED AS GUIDE RNA
2007: Barringou’s first experiment on CRISPR adaptive immunity found that spacer sequences were transcribed into guide RNAs.
August, 2008 — John van der Oost and colleagues at the University of Wageningen, Netherlands showed that in E. coli, spacer sequences, which are derived from phages are transcribed into small RNAs, termed CRISPR RNAs (crRNAs), that guide Cas proteins to the target DNA for double strand breakage of DNA (Brouns et al), 2008). CRISPR helps find the Viral DNA target.
CRISPR ACTS UPON DNA TARGETS NOT RNA
2008: Merraffini and Boutin confirmed that CRISPR acts upon DNA targets.
December, 2008 — Luciano Marraffini and Erik Sontheimer, Northwestern University, Illinois found that the target molecule is the DNA not RNA (often implicated in silencing mechanism). They hypothesized that CRISPR- Cas system could be a powerful tool applicable to non-bacterial DNAs including those of mammalians.
2009: Hale finds that the Type III-B Cmr complexes cleave RNA also in addition to Cas9.
CRISPR Cas9 CREATES DOUBLE STRAND DNA BREAK
December, 2010 — Sylvain Moineau, University of Laval, Quebec City, Canada reports that CRISPR-Cas9 creates double-stranded breaks in target DNA at precise positions, 3 nucleotides upstream of the PAM (Garneau et al., 2010). They also confirmed that Cas9 is the only protein required for cleavage in the CRISPR-Cas9 type II system, a distinguishing feature of Type II CRISPR systems.
TRACER RNA FORMS A DUPLEX STRUCTURE WITH crRNA IN ASSOCIATION WITH Cas9
March, 2011 — Emmanuelle Charpentier, Umea University, Sweden and University of Vienna, Austria discovered that in addition to the crRNA, a second small RNA exists, which they called trans-activating CRISPR RNA (tracrRNA). They showed that tracrRNA forms a duplex with crRNA, and that it is this duplex that guides Cas9 to its targets. Also, they showed that the CRISPR systems can function in other heterologous species.
2011: Garmelli reported that tracrRNA forms a duplex structure with crRNA in association with Cas9
CRISPR-Cas LOCUS CLONED
July, 2011 — Virginijus Siksnys of Vilnius University, Lithuania, cloned the entire CRISPR-Cas locus from S. thermophilus (a Type II system) and expressed it in E. coli (which does not contain a Type II system) and they demonstrated that it was capable of providing plasmid resistance (Sapranauskas et al., 2011). This suggested that CRISPR systems are self-contained units and verified that all of the required components of the Type II system were known. Biochemical characterization of Cas9-mediated cleavage was thus confirmed.
CRISPR REGION OF DNA IS TRANSCRIBED INTO crRNA THAT CAN ROAM ABOUT THE CELL AND TEAM UP WITH Cas9 ENDONULEASE PROTEINS AS MOLECULAR SCISSORS THAT CREATE DOUBLE STRAND DNA BREAK.
2011: Charpentier found trcrRNA for Cas9.
2012: Deticheva showed in vitro characterization of DNA targeting by Cas9.
CLEAVAGE SITE REQUIRES PAM (PROTOSPACER ACTIVATING MOTIF)
September, 2012 — Virginijus Siksnys and colleagues at the Vilnius University, Lithuania verified the cleavage site and the requirement for the PAM, and using point mutations, they further showed that the RuvC domain cleaves the non-complementary strand while the HNH domain cleaves the complementary site. They also noted that the crRNA could be trimmed down to a 20- nucleotide stretch sufficient for efficient cleavage. Most impressively, they showed that they could reprogram Cas9 to target a site of their choosing by changing the sequence of the crRNA.
crRNA and tracrRNA CAN BE FUSED TO FUNCTION AS SIGLE RNA
June, 2012 — Charpentier and Jennifer Doudna, University of California, Berkeley (2012) reported similar findings as those of Gasiunas et al. Charpentier and Doudna also reported that the crRNA and the tracrRNA could be fused together to create a single, synthetic guide, further simplifying the system (Although published in June 2012, this paper was submitted after Gasiunas et al.). CRISPR-Cas9 can now be harnessed for editing any genome and Professor Jennifer Doudna of UC Berkeley became the principal discoverer of the gene-editing tool CRISPR/Cas9.
Journals Science and PNAS (2012): The bacterial CRISPR-Cas9 chops up any DNA, not just that of viruses. The natural CRISPR system was transformed into a simple, programmable tool for genome editing.
FIRST APPLICATION IN EUKAYOTES
2013: Jinek, Cong, and Gushnas came with first application in Eukaryotes.
January, 2013 — Feng Zhang, Broad Institute of MIT and Harvard, McGovern Institute for Brain Research at MIT, Massachusetts was first to successfully adapt CRISPR-Cas9 for genome editing in eukaryotic cells (Cong et al., 2013). Zhang and his team engineered two different Cas9 orthologous genes in different species that evolved from a common ancestral gene by speciation (from S. thermophilus and S. pyogenes) and demonstrated targeted genome cleavage in human and mouse cells. They also showed that the system (i) could be programmed to target multiple genomic loci, and (ii) could drive homology-directed repair. Researchers from George Church’s lab at Harvard University reported similar findings in the same issue of Science (Mali et al., 2013).
RESOLVED CRYSTAL STRUCTURE OF Cas9-crRNA-tracrRNA COMPLEX FURTHER REVEALED BY HIGH SPEED ATOMIC FORCE MICROSCOPY
2014: Wang and Jinek performed genome wide functional screening with Cas9 and reported the crystal structure of Apo-Cas9 and the crystal structure of Cas9 in complex with RNA and Target DNA.
2017: Mikihiro Shibata etal. of the Kanazawa University in Japan and Hiroshi Nishimasu of the University of Tokyo visualized the CRISPR-Cas9 complex by high speed atomic force microscopy (Nature Communications, 10(8): 1430, 2017).
Dec 30, 2019” March, 2011 — Emmanuelle Charpentier, Umea University, Sweden and University of Vienna, Austria discovered that in addition to the crRNA, a second small RNA exists, which they called trans-activating CRISPR RNA (tracrRNA). They showed that tracrRNA forms a duplex with crRNA, and that it is this duplex that guides Cas9 to its targets. Also, they showed that the CRISPR systems can function in other heterologous species.
January 23, 2019: Grunwald etal. Use CRISPR-Cas for genetic control in Mice.
December 30, 2019: Jiankui in China is convicted for CRISPR-Cas9 use for creating human babies.
October 7, 2020: Drs. Emmanuelle Charpentier and Jennifer Doudna win Chemistry Nobel Prize for their work on chemical biology of the CRISPR- Cas system.
September 27, 2022: Vertex gets green light by FDA to treat sickle cell anemia.
Nov 10, 2022: Foy Ribas Mandle conducts small clinical trial about killing cancer cells by CRISPR editing.
Nov 23, 2022: Al Shayeb etal. Found thousands of phages by the CRISPR editing tool.
REFERENCES
1. Abbott TR, Dhamdhere G, Liu Y, Lin X, Goudy L, Zeng L, et al. (May 2020). “Development of CRISPR as an Antiviral Strategy to Combat SARS-CoV-2 and Influenza”. Cell. 181 (4): 865
2. S Barrangou, R. (2012). RNA-mediated programmable DNA cleavage. Nat. Biotechnol. 30, 836–838. Barrangou, R., and Marraffini, L.A. (2014). CRISPR-Cas systems: Prokaryotes upgrade to adaptive immunity. Mol. Cell 54, 234–244.
3. Barrangou, R., Fremaux, C., Deveau, H., Richards, M., Boyaval, P., Moineau, S., Romero, D.A., and Horvath, P. (2007). CRISPR provides acquired resistance against viruses in prokaryotes. Science 315, 1709–1712.
4. Bibikova, M., Carroll, D., Segal, D.J., Trautman, J.K., Smith, J., Kim, Y.G., and Chandrasegaran, S. (2001). Stimulation of homologous recombination through targeted cleavage by chimeric nucleases. Mol. Cell. Biol. 21, 289–297. 26 Cell 164, January 14, 2016 ª2016 Elsevier Inc.
5. Boch, J., Scholze, H., Schornack, S., Landgraf, A., Hahn, S., Kay, S., Lahaye, T., Nickstadt, A., and Bonas, U. (2009). Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326, 1509–1512.
6. Bolotin, A., Quinquis, B., Sorokin, A., and Ehrlich, S.D. (2005). Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin. Microbiology 151, 2551–2561.
7. Brouns, S.J.J., Jore, M.M., Lundgren, M., Westra, E.R., Slijkhuis, R.J.H., Snijders, A.P.L., Dickman, M.J., Makarova, K.S., Koonin, E.V., and van der Oost, J. (2008). Small CRISPR RNAs guide antiviral defense in prokaryotes. Science 321, 960–964.
8. Barrangou, R., Fremaux, C., Deveau, H., Richards, M., Boyaval, P., Moineau, S., Romero, D.A., and Horvath, P. (2007). CRISPR provides acquired resistance against viruses in prokaryotes. Science 315, 1709–1712.
9. Bolotin, A., Quinquis, B., Sorokin, A.,and Ehrlich, S.D. (2005). Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin. Microbiology 151, 2551–2561.
10. Brouns, S.J., Jore, M.M., Lundgren, M., Westra, E.R., Slijkhuis, R.J., Snijders, A.P., Dickman, M.J., Makarova, K.S., Koonin, E.V., van der Oost, J. (2008) Small CRISPR RNAs guide antiviral defense in prokaryotes. Science 321, 960-964.
11. Capecchi, M.R. (2005). Gene targeting in mice: functional analysis of the mammalian genome for the twenty-first century. Nat. Rev. Genet. 6, 507–512.
12. Carroll, D. (2012). A CRISPR approach to gene targeting. Mol. Ther. 20, 1658– 1660.
13. Cho, S.W., Kim, S., Kim, J.M., and Kim, J.-S. (2013). Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol. 31, 230–232.
14. Cong, L., Ran, F.A., Cox, D., Lin, S., Barretto, R., Habib, N., Hsu, P.D., Wu, X., Jiang, W., Marraffini, L.A., and Zhang, F. (2013). Multiplex genome engineering using CRISPR/Cas systems. Science 339, 819–823.
15. Chunyo, Liao and Chase, L. Baseil. 2021. The tracrRNA in CRISPR biology and technology, Annual Rev. Genetics, 55: 161-181
16. Daniel, Allen etal. 2021. Using synthetically engineered guide RNA to enhance CRISPR genpme editing systems in mammalian cells, Front Genome Ed, January 28.
17. Deltcheva, E., Chylinski, K., Sharma, C.M., Gonzales, K., Chao, Y., Pirzada, Z.A., Eckert, M.R., Vogel, J., and Charpentier, E. (2011). CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 471, 602–607.
18. Deveau, H., Barrangou, R., Garneau, J.E., Labonte´ , J., Fremaux, C., Boyaval, P., Romero, D.A., Horvath, P., and Moineau, S. (2008). Phage response to CRISPR-encoded resistance in Streptococcus thermophilus. J. Bacteriol. 190, 1390–1400.
19. Gasiunas, G., Barrangou, R., Horvath, P., and Siksnys, V. (2012). Cas9–crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Pnas 109, E2579–E2586.
20. Garneau, J.E., Dupuis, M.-E` ., Villion, M., Romero, D.A., Barrangou, R., Boyaval, P., Fremaux, C., Horvath, P., Magada´ n, A.H., and Moineau, S. (2010). The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 468, 67–71.
21. Haber, J.E. (2000). Lucky breaks: analysis of recombination in Saccharomyces. Mutat. Res. 451, 53–69.
22. Hale, C.R., Zhao, P., Olson, S., Duff, M.O., Graveley, B.R., Wells, L., Terns, R.M., and Terns, M.P. (2009). RNA-guided RNA cleavage by a CRISPR RNA-Cas protein complex. Cell 139, 945–956.
23. Horvath, P., Romero, D.A., Couˆ te´-Monvoisin, A.-C., Richards, M., Deveau, H., Moineau, S., Boyaval, P., Fremaux, C., and Barrangou, R. (2008). Diversity, activity, and evolution of CRISPR loci in Streptococcus thermophilus. J. Bacteriol. 190, 1401–1412.
24. Hsu, P.D., Lander, E.S., and Zhang, F. (2014). Development and applications of CRISPR-Cas9 for genome engineering. Cell 157, 1262–1278.
25. Ishino, Y., Shinagawa, H., Makino, K., Amemura, M., and Nakata, A. (1987). Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 169, 5429–5433.
26. Jansen, R., Embden, J.D.A.V., Gaastra, W., and Schouls, L.M. (2002). Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 43, 1565–1575.
27. Jasin, M., and Rothstein, R. (2013). Repair of strand breaks by homologous recombination. Cold Spring Harb. Perspect. Biol. 5, a012740.
28. Jiang, W., and Marraffini, L.A. (2015). CRISPR-Cas: New tools for genetic manipulations from bacterial immunity systems. Annu. Rev. Microbiol. 69, 209–228.
29. Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J.A., and Charpentier, E. (2012). A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816–821.
30. Jinek, M., East, A., Cheng, A., Lin, S., Ma, E., and Doudna, J. (2013). RNA-programmed genome editing in human cells. eLife 2, e00471.
31. Kennedy EM, Cullen BR (May 2015). “Bacterial CRISPR/Cas DNA endonucleases: A revolutionary technology that could dramatically impact viral research and treatment”. Virology. 479–480: 213–20
32. Makarova, K.S., Grishin, N.V., Shabalina, S.A., Wolf, Y.I., Koonin, E.V. (2006). A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action. Biology Direct 2006, 1:7.
33. Mali, P., Yang, L., Esvelt, K.M., Aach, J., Guell, M., DiCarlo, J.E., Norville, J.E., and Church, G.M. (2013). RNA-guided human genome engineering via Cas9. Science 339, 823–826.
34. Marraffini, L.A., and Sontheimer, E.J. (2008). CRISPR interference limits horizontal gene transfer in staphylococci by targeting DNA. Science 322, 1843–1845.
35. Makarova, K.S., Grishin, N.V., Shabalina, S.A., Wolf, Y.I., and Koonin, E.V. (2006). A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action. Biol. Direct 1, 7.
36. Makarova, K.S., Wolf, Y.I., Alkhnbashi, O.S., Costa, F., Shah, S.A., Saunders, S.J., Barrangou, R., Brouns, S.J.J., Charpentier, E., Haft, D.H., et al. (2015). An updated evolutionary classification of CRISPR-Cas systems. Nat. Rev. Microbiol. 13, 722–736.
37. Mangold, M., Siller, M., Roppenser, B., Vlaminckx, B.J.M., Penfound, T.A., Klein, R., Novak, R., Novick, R.P., and Charpentier, E. (2004). Synthesis of group A streptococcal virulence factors is controlled by a regulatory RNA molecule. Mol. Microbiol. 53, 1515–1527.
38. Medawar, P. (1968). Lucky Jim (New York Review of Books), March 28, 1968.
39. Miller, J.C., Tan, S., Qiao, G., Barlow, K.A., Wang, J., Xia, D.F., Meng, X., Paschon, D.E., Leung, E., Hinkley, S.J., et al. (2011). A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 29, 143–148.
40. Mojica, F.J.M., and Garrett, R.A. (2012). Discovery and Seminal Developments in the CRISPR Field. In CRISPR-Cas Systems, R. Barrangou and J. van der Oost, eds. (Berlin, Heidelberg: Springer Berlin Heidelberg), pp. 1–31.
41. Mojica, F.J.M., D ez-Villase or, C.S., Garc a-Mart nez, J.S., and Soria, E. (2005). Intervening Sequences of Regularly Spaced Prokaryotic Repeats Derive from Foreign Genetic Elements. J Mol Evol 60, 174–182.
42. Mojica, F.J.M., Juez, G., and Rodrı´guez-Valera, F. (1993). Transcription at different salinities of Haloferax mediterranei sequences adjacent to partially modified PstI sites. Mol. Microbiol. 9, 613–621.
43. Mojica, F.J.M., Ferrer, C., Juez, G., and Rodrı´guez-Valera, F. (1995). Long stretches of short tandem repeats are present in the largest replicons of the Archaea Haloferax mediterranei and Haloferax volcanii and could be involved in replicon partitioning. Mol. Microbiol. 17, 85–93.
44. Mojica, F.J.M., Dı´ez-Villasen˜ or, C., Soria, E., and Juez, G. (2000). Biological significance of a family of regularly spaced repeats in the genomes of Archaea, Bacteria and mitochondria. Mol. Microbiol. 36, 244–246.
45. Mojica, F.J.M., Dı´ez-Villasen˜ or, C., Garcı´a-Martı´nez, J., and Soria, E. (2005). Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. J. Mol. Evol. 60, 174–182.
46. Nishimasu H. etal. 2018 Crystal structure of Cas9-NG, Science, 361: 1259-1262
47. Nishimasu, Hiroshi. 2014. Crystal structure of Cas9 in complex with gRNA and target DNA, Cell, 156 (5): 935-949
48. Pandika, M. (2014) Jennifer Doudna, CRISPR Code Killer, Ozy.com, January 7, 2014. < http://www.ozy.com/rising-stars/jennifer-doudna-crispr-codekiller/4690 >.
49. Porteus, M.H., and Baltimore, D. (2003). Chimeric nucleases stimulate gene targeting in human cells. Science 300, 763.
50. Pourcel, C., Salvignol, G., and Vergnaud, G. (2005). CRISPR elements in Yersinia pestis acquire new repeats by preferential uptake of bacteriophage DNA, and provide additional tools for evolutionary studies. Microbiology 151, 653–663.
51. Sapranauskas, R., Gasiunas, G., Fremaux, C., Barrangou, R., Horvath, P., and Siksnys, V. (2011). The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli. Nucl. Acids Res. 39, gkr606–gkr9282.
52. Sander, J.D., and Joung, J.K. (2014). CRISPR-Cas systems for editing, regulating and targeting genomes. Nat. Biotechnol. 32, 347–355. Cell 164, January 14, 2016 ª2016 Elsevier Inc. 27
53. Sapranauskas, R., Gasiunas, G., Fremaux, C., Barrangou, R., Horvath, P., and Siksnys, V. (2011). The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli. Nucleic Acids Res. 39, 9275–9282.
54. Shah Shiraz A etal. 20132. Protospacer recognition motifs, RNA Biol., 10(5): 889-899
55. Sharma, C.M., Hoffmann, S., Darfeuille, F., Reignier, J., Findeiss, S., Sittka, A., Chabas, S., Reiche, K., Hackermu¨ ller, J., Reinhardt, R., et al. (2010). The primary transcriptome of the major human pathogen Helicobacter pylori. Nature 464, 250–255.
56. Siksnys, V., Gasiunas, G., and Karvelis, T. (2012). RNA-directed DNA cleavage by the Cas9-crRNA complex from CRISPR3/Cas immune system of Streptococcus thermophilus. U.S. Provisional Patent Application 61/613,373, filed March 20, 2012; later published as US2015/0045546 (pending).
57. Sontheimer, E., and Marraffini, L. (2008). Target DNA interference with crRNA. U.S. Provisional Patent Application 61/009,317, filed September 23, 2008; later published as US2010/0076057 (abandoned).
58. Sorek, R., Kunin, V., and Hugenholtz, P. (2008). CRISPR–a widespread system that provides acquired resistance against phages in bacteria and archaea. Nat. Rev. Microbiol. 6, 181–186.
59. Sternberg, S.H., and Doudna, J.A. (2015). Expanding the Biologist’s Toolkit with CRISPR-Cas9. Mol. Cell 58, 568–574. Travis, J. (2015). GENETIC ENGINEERING. Germline editing dominates DNA summit. Science 350, 1299–1300.
60. Tristan, Scott. 2019. Improved Cas9 activity by by modification of the tracrRNA, Scientific Reports, Article No. 16104
61. Westra, E.R., Jackson, R.N., and Wiedenheft, B. (2014). Unravelling the structural and mechanistic basis of CRISPR-Cas systems. Nat. Rev. Microbiol. 12, 479–492.
62. Wright, A.V., James, K., Nun˜ ez, J.K., and Doudna, J.A. (2016). Biology and applications of CRISPR systems: Harnessing nature’s toolbox for genome engineering. Cell 164, this issue, 29–44.
63. Yamado, Mari etal. 2017. Crystal structure of the minimal Cas9 for camphylobacter jejuni reveal the molecular diversity in the CRISPR Cas9 system, Molecular Cell, 65: 1109-1121
64. Zetsche, B., Gootenberg, J.S., Abudayyeh, O.O., Slaymaker, I.M., Makarova, K.S., Essletzbichler, P., Volz, S.E., Joung, J., van der Oost, J., Regev, A., et al. (2015). Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell.
65. Zhang, F. (2012). Systems Methods and Compositions for Sequence Manipulation. U.S. Provisional Patent Application 61/736,527, filed December 12, 2012; later published as US008697359B1 (awarded).
Leave a Reply